
Les chercheurs sont longtemps passés à côté de l’idée que les humains pouvaient être de bons coureurs d’endurance. Jusqu’au début des années 1980, il était couramment admis que la bipédie avait donné aux humains primitif de grands avantages via la marche, mais que nous étions de piètres coureurs, notamment parce que l’on s’arrêtait au constat de nos faibles qualités de sprinteurs.
Mais dans les années 1980, un chercheur, lui-même marathonien, David Carrier, a émis l’hypothèse que nos ancêtres aient pu être d’excellents coureurs d’endurance, et que cette qualité aurait pu les aider en permettant la pratique d’un type de chasse particulier, appelé chasse à l’épuisement. Passée pratiquement inaperçue, l’idée a été reprise une vingtaine d’années plus tard, si l’idée d’une chasse à l’épuisement pure ne fait pas l’unanimité, les capacités d’endurance humaine exceptionnelles pour une espèce de primates ont très probablement eu un rôle important dans l’évolution humaine.
Par ailleurs, l’idée que les premiers coureurs aient couru nus-pieds laisse entendre que la technique de course naturelle des humains est différente de la technique utilisée par les coureurs chaussés (attaque sur le milieu du pied pour la technique naturelle, sur le talon pour les coureurs chaussés), et pourrait expliquer que ces derniers se blessent souvent. La question reste très débattue, et l’intérêt d’apprendre à courir pieds nus pour retrouver une foulée naturelle est âprement discuté.
1. Adaptations à la course d’endurance
2. Implications modernes
[Retour au sommaire des données scientifiques]
Adaptations à la course d’endurance
Ethnography and ethnohistory support the efficiency of hunting through endurance running in humans [Abstract]
Morin & Winterhalder
Nature Human Behaviour, 2024
We likewise analyse an ethnohistoric and ethnographic database of nearly 400 EP cases representing 272 globally distributed locations. We provide estimates for return rates of EPs and argue that these are comparable to other pre-modern hunting methods in specified contexts. EP hunting as a method of food procurement would have probably been available and attractive to Plio/Pleistocene hominins.
–
Are humans evolved specialists for running in the heat? Man vs. horse races provide empirical insights [Texte] [PDF]
Halsey & Bryce
Experimental physiology, 2020
Comparing the results of races that pit man against horse, we find that ambient temperature on race day has less deleterious effects on running speed in humans than it does on their quadrupedal adversary. This is evidence that humans are adapted for endurance running at high ambient temperatures. We debate whether this supports the hypothesis that early man was evolutionarily adapted for persistence hunting.
Combien de temps un Homo erectus était-il capable de courir sans boire ? Environ 5h, selon cette étude :
Dehydration and persistence hunting in Homo erectus [Texte]
Martin Hora et al.
Journal of human evolution, 2020
Our results showed that H. erectus would reach the dehydration limit in 5.5–5.7 h of persistence hunting at the reported Kalahari conditions, which we argue represent a conservative model also for Early Pleistocene East Africa. Maximum hunt duration without drinking was negatively related to the relative body surface area of the hunter. Moreover, H. erectus would be able to persistence hunt over 5 h without drinking despite possible deviations from modern-like heat dissipation capacity, aerobic capacity, and locomotor economy. We conclude that H. erectus could persistence hunt large prey without the need to carry water.
–
Rethinking the evolution of the human foot: insights from experimental research [PDF]
Holowka & Lieberman
Journal of experimental biology, 2018
We use this framework to interpret the fossil record and argue that the human foot passed through three evolutionary stages: first, a great ape-like foot adapted for arboreal locomotion but with some adaptations for bipedal walking; second, a foot adapted for effective bipedal walking but retaining some arboreal grasping adaptations; and third, a human-like foot adapted for enhanced economy during long-distance walking and running that had lost its prehensility. Based on this scenario, we suggest that selection for bipedal running played a major role in the loss of arboreal adaptations.
–
L’inactivation d’un gène il y a deux ou trois millions d’années a pu améliorer les capacités d’endurances de nos ancêtres.
Human-like Cmah inactivation in mice increases running endurance and decreases muscle fatigability: implications for human evolution [PDF]
Okerblom et al.
Proceedings of the royal society B, 2018
Taken together, these data suggest that CMAH loss contributes to an improved skeletal muscle capacity for oxygen use. If translatable to humans, CMAH loss could have provided a selective advantage for ancestral Homo during the transition from forest dwelling to increased resource exploration and hunter/gatherer behaviour in the open savannah.
Economy and Endurance in Human Evolution [Texte]
Herman Pontzer
Current biology, 2017
The appearance of skeletal traits related to endurance (e.g., larger limb joints, spring-like plantar arch) in Homo was somewhat mosaic, with the full endurance suite apparent only ∼1 million years ago. The development of endurance capabilities in Homo appears to parallel the evolutionary increase in brain size, cognitive sophistication, and metabolic rate.
Evolutionary Implications of Persistence Hunting: An Examination of Energy Return on Investment for !Kung Hunting [PDF]
Glaub & Hall
Human ecology, 2017
We estimate that the EROI of these hunter-gatherers hunting a kudu ranged from 26:1 to 69:1. The net energy gained from such an effort would sustain an average sized !Kung family for 6.7 to 11.2 days. The “profit” energy within these ranges would have supported the early human societies that practiced persistence hunting, contributing to the often-noted “leisure” characterizing many foraging societies
These bones where made for jogging : an analysis of the lower limb skeletal evidence for the endurance running hypothesis [PDF]
Kim P. Deckers
Inter-section, 2017
A complete suite of anatomical features related to endurance running is not seen until Homo ergaster at 1.8 million years ago. However, a mosaic of these features is present in earlier Australopiths. This, combined with the lack of evidence for long range projectile weapons in the Pleistocene, indicates these features may have been positively selected for to increase walking efficiency and increased endurance running capabilities were just a fortunate by-product. Currently, there is insufficient evidence to support the notion that anatomical changes observed in early Homo evolved to increase running capabilities.
Investigating the case of human nose shape and climate adaptation
Zaidi et al.
Plos Genetics, 2017
The evolutionary reasons for variation in nose shape across human populations have been subject to continuing debate. An import(ant ?) function of the nose and nasal cavity is to condition inspired air before it reaches the lower respiratory tract.
Additive opportunistic capture explains group hunting benefits in African wild dogs [Texte]
Hubel et al.
Nature communications, 2016
African wild dogs (Lycaon pictus) are described as highly collaborative endurance pursuit hunters based on observations derived primarily from the grass plains of East Africa. However, the remaining population of this endangered species mainly occupies mixed woodland savannah where hunting strategies appear to differ from those previously described.
–
Human Locomotion and Heat Loss: An Evolutionary Perspective [PDF]
Daniel Lieberman
Comprehensive physiology, 2015
Humans are unique in many respects including being furless, striding bipeds that excel at walking and running long distances in hot conditions. This review summarizes what we do and do not
know about the evolution of these characteristics, and how they are related. Although many details remain poorly known, the first hominins (species more closely related to humans than to
chimpanzees) apparently diverged from the chimpanzee lineage because of selection for bipedal walking, probably because it improved their ability to forage efficiently. However, because bipedal hominins are necessarily slow runners, early hominins in open habitats likely benefited from improved abilities to dump heat in order to forage safely during times of peak heat when predators
were unable to hunt them. Endurance running capabilities evolved later, probably as adaptations for scavenging and then hunting. If so, then there would have been strong selection for heat-loss mechanisms, especially sweating, to persistence hunt, in which hunters combine endurance running and tracking to drive their prey into hyperthermia. As modern humans dispersed into a wide range of habitats over the last few hundred thousand years, recent selection has helped populations cope better with a broader range of locomotor and thermoregulatory challenges, but all humans remain essentially adapted for long distance locomotion rather than speed, and to dump rather than retain heat.
–
Prey mortality profiles indicate that Early Pleistocene Homo at Olduvai was an ambush predator [Abstract]
Bunn & Gurtov
Quaternary international, 2014
The prime-dominated profile at FLK Zinj is significantly different from profiles formed by the three scavenging methods, which likely indicates hunting by Early Pleistocene Homo.
D’extraordinaires capacités de course d’endurance n’ont probablement pas été très utiles pour le charognage.
Endurance running and its relevance to scavenging by early hominids
Ruxton & Wilkinson
Evolution, 2013
Our model estimates that nonhominin scavengers will generally be able to reach the carcass within 30 min of detecting a plume of vultures above a nearby carcass. We argue that endurance running over periods greater than 30 min would not have provided a selective advantage to early hominins through increased scavenging opportunities. However, shorter distance running may have been selected, particularly if hominins could defend or usurp carcasses from other mammalian scavengers.
Thermoregulation and endurance running in extinct hominins: Wheeler’s models revisited [Texte]
Ruxton & Wilkinson
Journal of human evolution, 2011
Our model suggests that for endurance running to be possible, a hominin would need locomotive efficiency, sweating rates, and areas of hairless skin similar to modern humans. We argue that these restrictions suggest that endurance running may have been possible (from a thermoregulatory viewpoint) for Homo erectus, but is unlikely for any earlier hominins.
Bovid mortality profiles in paleoecological context falsify hypotheses of endurance running–hunting and passive scavenging by early Pleistocene hominins [Abstract]
Bunn & Pickering
Quaternary research, 2010
In all but one case, the bovid mortality data from FLK Zinj violate test predictions of the endurance running-hunting and passive scavenging hypotheses. When combined with other taphonomic data, these results falsify both hypotheses, and lead to the hypothesis that early Homo operated successfully as an ambush predator.
Optimal running speed and the evolution of hominin hunting strategies [PDF]
Steudel-Numbers & Wall-Scheffler
Journal of human evolution, 2009
The belief in the lack of optimal running speeds in humans is based, however, on a number of early studies with experimental designs inadequate for the purpose of evaluating optimality. Here we measured the energetic cost of human running (n = 9) at six different speeds for five minutes at each speed, with careful replicates and controls. We then compared the fit of linear versus curvilinear models to the data within each subject. We found that individual humans do, in fact, have speeds at which running is significantly less costly than at other speeds (i.e., an optimal running speed). In addition, we demonstrate that the use of persistence hunting methods to gain access to prey at any running speed, even the optimum, would be extremely costly energetically, more so than a persistence hunt at optimal walking speed. We argue that neither extinct nor extant hominin populations are as flexible in the chosen speeds of persistence hunting pursuits as other researchers have suggested.
Il a été argumenté que les chasseurs-cueilleurs contemporains ne pratiquent pas la chasse à l’épuisement. Il semble difficile d’en tirer des conclusions sur le comportement des premiers humains, avant l’invention d’armes sophistiquées ou la domestication du chien et du cheval. De plus, l’absence d’observation de chasses à l’épuisement de la part des ethnologues ne reflète peut-être pas l’incapacité des chasseurs cueilleurs à mener ces chasses, mais seulement l’incapacité des ethnologues à les documenter (note : quelques cas de chasse à l’épuisement ont été tout de même observés, l’un d’eux a même été filmé).
The endurance running hypothesis and hunting and scavengingin savanna-woodlands [PDF]
Pickering & Bunn
Journal of human evolution, 2007
We agree with the opinion of Bramble and Lieberman(2004)that early Homo required a high-quality diet, which included a substantial meat component, and that it was thus a capable carcass forager. However, our understanding of the paleoenvironment, paleoecology, and archaeology of early Homo sites, reviewed here, makes us dubious about their further suggestion that ER might have been employed regularly and successfully in service of that foraging pursuit. We are re-luctant to assign to early Homo the impressive tracking skills of the Kalahari San, when the cognitive and meat-foraging abilities of Plio-Pleistocene Homo are active research issues.The behavioral pattern that selected for ER in the genus Homo remains unclear, but it seems likely that hunting and scavenging contributed minimally, if at all.
The Evolution of Endurance Running and the Tyranny of Ethnography: A Reply to Pickering and Bunn (2007) [PDF]
Daniel Lieberman et al.
Journal of human evolution, 2007
We will never know for sure why and how ER capabilities
evolved, but the modern ethnographic record is a limited, biased,
and sometimes misleading source of evidence to test hypotheses
about how ESA hominids hunted and scavenged.
The challenge for paleoanthropologists is to explain the past
in terms of testable hypotheses derived from actualistic studies
and middle-range research without succumbing to the ‘‘tyranny
of ethnography.’’
The relevance of persistence hunting to human evolution [PDF]
Louis Liebenberg
Journal of human evolution, 2008
As pointed out by Lieberman et al. (2007), Pickering and Bunn (2007) made several flawed assumptions. Persistence hunting may have been more common before the invention of the bow-and-arrow or the domestication of dogs and horses. The apparent scarcity of ethnographic records of PH does not imply that PH was rare it could simply be that anthropologists who were able to observe PH were rare. Over the last 50 years, hunter-gatherers in the Kalahari have experienced drastic changes, and the recent observations of PH may simply represent the tail-end of a dying tradition. In the absence of a better hypothesis, PH and scavenging remain plausible explanations for the evolution of ER.
The evolution of marathon running : capabilities in humans [PDF]
Lieberman & Bramble
Sports medicine, 2007
Humans have exceptional capabilities to run long distances in hot, arid conditions. These abilities, unique among primates and rare among mammals,derive from a suite of specialised features that permit running humans to store and release energy effectively in the lower limb, help keep the body’s center of mass stable and overcome the thermoregulatory challenges of long distance running. Human endurance running performance capabilities compare favourably with those of other mammals and probably emerged sometime around 2 million years ago in order to help meat-eating hominids compete with other carnivores.
Persistence hunting by modern hunter-gatherers [PDF]
Louis Liebenberg
Current anthropology, 2006
Endurance running may be a derived capability of the genus Homo and may have been instrumental in the evolution of the human body form. Two hypotheses have been presented to explain why early Homo would have needed to run long distances: scavenging and persistence hunting. Persistence hunting takes place during the hottest time of the day and involves chasing an animal until it is run to exhaustion. A critical factor is the fact that humans can keep their bodies cool by sweating while running. Another critical factor is the ability to track down an animal. Endurance running may have had adaptive value not only in scavenging but also in persistence hunting. Before the domestication of dogs, persistence hunting may have been one of the most efficient forms of hunting and may therefore have been crucial in the evolution of humans.
Le grand fessier, muscle essentiel dans notre évolution. Il semble avoir une utilité modeste pour la marche, mais beaucoup plus importante pour la course.
The human gluteus maximus and its role in running [PDF]
Daniel Lieberman et al.
Experimental Biology, 2006
The human gluteus maximus is a distinctive muscle in terms of size, anatomy and function compared to apes and other non-human primates. Here we employ electromyographic and kinematic analyses of human subjects to test the hypothesis that the human gluteus maximus plays a more important role in running than walking. The results indicate that the gluteus maximus is mostly quiescent with low levels of activity during level and uphill walking, but increases substantially in activity and alters its timing with respect to speed during running. The major functions of the gluteus maximus during running are to control flexion of the trunk on the stanceside and to decelerate the swing leg; contractions of the stance-side gluteus maximus may also help to control flexion of the hip and to extend the thigh. Evidence for when the gluteus maximus became enlarged in human evolution is equivocal, but the muscle’s minimal functional role during walking supports the hypothesis that enlargement of the gluteus maximus was likely important in the evolution of hominid running capabilities.
Article considéré comme fondateur dans la théorie de l’homme coureur d’endurance et chasseur à l’épuisement. Il reprend l’essentiel des travaux des années 1980 de David Carrier, en y ajoutant toutefois des considérations physiologiques importantes.
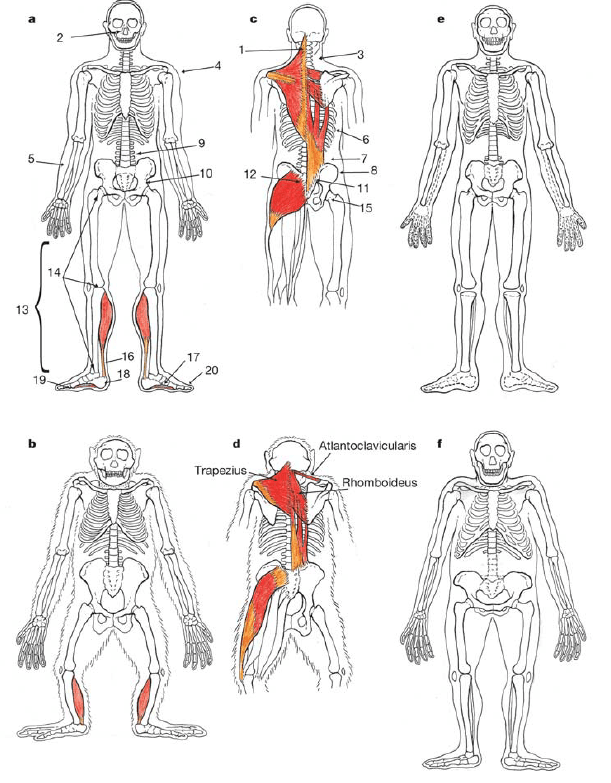
The loss of functional body hair in man: the influence of thermal environment, body form and bipedality
P.E. Wheeler
Journal of human evolution, 1985
An insulating layer of body hair is crucial to the thermoregulatory energetics of most mammals. Only some highly specialized aquatic forms and a few fossorial and terrestrial species which experience special heat stress problems have evolved a naked skin. Man’s body size and form suggest that his naked skin is associated with the prevention of hyperthermia in hot climates rather than an aquatic ancestry. The reduced direct solar radiation exposure of hominids in open equatorial environments, resulting from their unique bipedal posture, may explain the absence of this characteristic among other savannah mammals of comparable body mass.
The energetic paradox of human running and hominid evolution [PDF]
David Carrier
Current anthropology, 1984
This analysis of the morphology and physiology of human locomotion, although incomplete and largely speculative, sug- gests that man is a primate that has become specialized for (among other things) endurance running […] . Exercising anthropoid primates can dissipate heat more rapidly than certain cursorial mammals. This suggests that the earliest hominids may have been more effective at heat loss than many other contemporary mammals. Furthermore, early australo- pithecines were fully bipedal. As a result, they were probably not constrained to the typical 1:1 breathing pattern of running quadrupeds. Consequently, it is possible that one of the impor- tant factors which differentiated hominids from other an- thropoids, and ultimately led to the evolution of H. sapiens, was the occupation of a new predatory niche. This hypothetical niche was that of a diurnal predator which depended upon exceptional endurance in hot (midday) temperatures to disable swifter prey animals.
Le tout premier article suggérant des capacités d’endurances particulièrement développées chez les êtres humains. La bipédie permet de libérer les poumons des contraintes liées à la course (principalement au galop). Les humains ont donc la capacité de découpler leur rythme de course de leur respiration, et donc de maintenir très longtemps leur vitesse de course, là où les animaux ont besoin, à partir d’une certaine vitesse, de s’arrêter pour respirer.
Running and breathing in mammals [PDF]
Bramble & Carrier
Science, 1983
Quadrupedal species normally synchronize the locomotor and respiratory cycles at a constant ratio of 1:1 (strides per breath) in both the trot and gallop. Human runners differ from quadrupeds in that while running they employ several phase-locked patterns (4:1, 3:1, 2:1, 1:1, 5:2, and 3:2), although a 2:1 coupling ratio appears to be favored.
La forme du nez peut aussi avoir été utile à réguler la température de l’air inspiré. Elle change dans le même laps de temps que les autres modifications anatomiques liées à la course.
Nasal Morphology and the Emergence of Homo erectus [Abstract]
Franciscus & Trinkaus
American journal of physical anthropology, 1988
Modern humans, among extant hominoids, possess a unique projecting, external nose whose basic structure is reflected in a series of skeletal features including nasal bone convexity, an internasal angle, lateral nasal aperture eversion, prominence and anterior positioning of the anterior nasal spine, an acute angle of the subnasal alveolar clivus, and an expansion of the breadth of the nasal bones relative to that of the piriform aperture. This anatomy appears with the emergence of Homo erectus ca. 1.6 million years ago. Although it undoubtedly evolved in the context of craniofacial and dental reduction during hominid evolution, it appears to have been primarily a response to the need for moisture conservation in an arid environment via turbulence enhancement and ambient cooling of expired air. Its appearance at this time in hominid evolution, in conjunction with the presence of a fatigue- resistant locomotor anatomy characteristic of archaic members of the genus Homo, indicates a shift to increasingly prolonged bouts of activity in open and arid environments.
Implications modernes
https://lib.dr.iastate.edu/cgi/viewcontent.cgi?article=5662&context=etd
Une étude qui trouve que l’attaque talon est plus économe que l’attaque médio-pied, du moins pour les vitesses relativement faibles.
Rearfoot striking runners are more economical than midfoot strikers
Ana Ogueta-Alday et al.
Medecine & science in sport & exercise, 2014
https://www.researchgate.net/publication/256436455_Rearfoot_Striking_Runners_Are_More_Economical_Than_Midfoot_Strikers
Medecine & science in sport & exercise, 2012
https://pdfs.semanticscholar.org/5632/9749cfe1243f206a6c4fe4919c14253a9072.pdf
Et un peu de littérature, un livre que l’on ne saurait trop recommander (même s’il prend quelque libertés avec les questions scientifiques) :
Christopher McDougall, Born to run, éditions Gérin, 2012.

–
