
1. Forêt, déforestation, reforestation
2. Sujets d’écologie divers
Forêt, déforestation, reforestation
Global priority areas for ecosystem restoration
Strassburg et al.
Nature, 2020
https://www.nature.com/articles/s41586-020-2784-9
We find that restoring 15% of converted lands in priority areas could avoid 60% of expected extinctions while sequestering 299 gigatonnes of CO2—30% of the total CO2 increase in the atmosphere since the Industrial Revolution. The inclusion of several biomes is key to achieving multiple benefits. Cost effectiveness can increase up to 13-fold when spatial allocation is optimized using our multicriteria approach, which highlights the importance of spatial planning. Our results confirm the vast potential contributions of restoration to addressing global challenges, while underscoring the necessity of pursuing these goals synergistically.

–
Tree planting in organic soils does not result in net carbon sequestration on decadal timescales
Friggens et al.
Global change biology, 2020
https://onlinelibrary.wiley.com/doi/10.1111/gcb.15229
At all four experimental sites sampled, there was no net gain in ecosystem C stocks 12–39 years after afforestation—indeed we found a net ecosystem C loss in one of four sites with deciduous B. pubescens stands; no net gain in ecosystem C at three sites planted with B. pubescens; and no net gain at additional stands of P. sylvestris. We hypothesize that altered mycorrhizal communities and autotrophic C inputs have led to positive ‘priming’ of soil organic matter, resulting in SOC loss, constraining the benefits of tree planting for ecosystem C sequestration.
–
Assessing the ecological vulnerability of forest landscape to agricultural frontier expansion in the Central Highlands of Vietnam
Bourgoin et al.
International journal of applied earth observation and geoinformation, 2020
https://www.sciencedirect.com/science/article/pii/S0303243419307202
We map current forest-agricultural mosaics with high accuracy to assess landscape intensification (kappa index = 0.78). We also map the expansion of the agricultural frontier and highlighted heterogeneous agricultural encroachment on forested areas (kappa index = 0.72-0.93). Finally, we identify degradation and fragmentation trajectories that affect forest cover at different rates and intensity. Combined, these indicators pinpoint hotspots of forest vulnerability. This study provides tailored management responses and levers for action by local decision makers. The accessibility of multi-dimensional remote sensing data and the developed landscape approach open promising perspectives for continuously monitoring agricultural frontiers.
–
Vegetation structural change since 1981 significantly enhanced the terrestrial carbon sink
Chen et al.
Nature communication, 2019
https://www.nature.com/articles/s41467-019-12257-8

–
The global tree restoration potential
Bastin et al.
Science, 2019
https://science.sciencemag.org/content/365/6448/76
The restoration of trees remains among the most effective strategies for climate change mitigation. We mapped the global potential tree coverage to show that 4.4 billion hectares of canopy cover could exist under the current climate. Excluding existing trees and agricultural and urban areas, we found that there is room for an extra 0.9 billion hectares of canopy cover, which could store 205 gigatonnes of carbon in areas that would naturally support woodlands and forests. This highlights global tree restoration as one of the most effective carbon drawdown solutions to date. However, climate change will alter this potential tree coverage. We estimate that if we cannot deviate from the current trajectory, the global potential canopy cover may shrink by ~223 million hectares by 2050, with the vast majority of losses occurring in the tropics. Our results highlight the opportunity of climate change mitigation through global tree restoration but also the urgent need for action.
–
Satellite-observed pantropical carbon dynamics
Lei Fan et al.
Nature plants, 2019
https://www.nature.com/articles/s41477-019-0478-9
Here, we used low-frequency L-band passive microwave observations to compute a direct and spatially explicit quantification of annual aboveground carbon (AGC) fluxes and show that the tropical net AGC budget was approximately in balance during 2010 to 2017, the net budget being composed of gross losses of −2.86 PgC yr−1 offset by gross gains of −2.97 PgC yr−1 between continents. Large interannual and spatial fluctuations of tropical AGC were quantified during the wet 2011 La Niña year and throughout the extreme dry and warm 2015–2016 El Niño episode. These interannual fluctuations, controlled predominantly by semiarid biomes, were shown to be closely related to independent global atmospheric CO2 growth-rate anomalies (Pearson’s r = 0.86), highlighting the pivotal role of tropical AGC in the global carbon budget.
–
Grasslands may be more reliable carbon sinks than forests in California
Dass et al.
Environmental reserch letter, 2018
https://iopscience.iop.org/article/10.1088/1748-9326/aacb39/meta
Using a set of modeling experiments, we show that California grasslands are a more resilient C sink than forests in response to 21st century changes in climate, with implications for designing climate-smart Cap and Trade offset policies. The resilience of grasslands to rising temperatures, drought and fire, coupled with the preferential banking of C to belowground sinks, helps to preserve sequestered terrestrial C and prevent it from re-entering the atmosphere. In contrast, California forests appear unable to cope with unmitigated global changes in the climate, switching from substantial C sinks to C sources by at least the mid-21st century.
–
Corridors best facilitate functional connectivity across a protected area network
Stewart et al.
Nature scientific reports, 2018
Cliquer pour accéder à s41598-019-47067-x.pdf
Using a tractable PA network mesocosm, we employ cutting-edge biologging technology to analyze animal movement decisions in response to a highly heterogeneous landscape. We analyze these data to test, in a novel way, three common hypotheses about functional connectivity – structural corridors, least cost paths, and stepping stones. Consistently, animals moved along structurally self-similar corridors. In reference to the Aichi 2020 Biodiversity Targets, relying on species to “stepping stone” across habitat remnants may not achieve protected area network conservation objectives
–
Although aggressive emission control strategies have been implemented recently in the Beijing–Tianjin–Hebei area (BTH), China, pervasive and persistent haze still frequently engulfs the region during wintertime. Afforestation in BTH, primarily concentrated in the Taihang and Yan Mountains, has constituted one of the controversial factors exacerbating the haze pollution due to its slowdown of the surface wind speed. We report here an increasing trend of forest cover in BTH during 2001–2013 based on long-term satellite measurements and the impact of the afforestation on the fine-particle (PM2.5) level. Simulations using the Weather Research and Forecast model with chemistry reveal that afforestation in BTH since 2001 has generally been deteriorating the haze pollution in BTH to some degree, enhancing PM2.5 concentrations by up to 6 % on average. Complete afforestation or deforestation in the Taihang and Yan Mountains would increase or decrease the PM2.5 level within 15 % in BTH. Our model results also suggest that implementing a large ventilation corridor system would not be effective or beneficial to mitigate the haze pollution in Beijing.
–
The importance of scattered trees for biodiversity conservation: A global meta‐analysis
Prevedello et al.
Journal of applied ecology, 2017
https://besjournals.onlinelibrary.wiley.com/doi/abs/10.1111/1365-2664.12943%4010.1111/%28ISSN%291365-2664.TropicalDiversity?casa_token=uEUsRpHYEvoAAAAA:Wj3q0f5CwthfHe2ElusHR4unXnflp4tOdXHSduKZ6qdLDnrPfTJ-bEOgcxi1c9GisnYRfkyRRof0NUOD
Areas with scattered trees support greater levels of biodiversity than open areas, as well as communities that are more similar to those inhabiting habitat patches. Scattered trees can be regarded as keystone structures for vertebrates, arthropods and terrestrial plants in landscapes world‐wide. The maintenance of scattered trees may be compatible with livestock grazing in some agricultural landscapes. Greater management effort and targeted, long‐term policies are needed to retain or re‐establish scattered trees in many farming landscapes in both forest and non‐forest biomes around the world.
–
The biodiversity cost of carbon sequestration in tropical savanna
Abreu et al.
Science advances, 2017
https://advances.sciencemag.org/content/3/8/e1701284
Fire suppression resulted in increased carbon stocks of 1.2 Mg ha−1 year−1 since 1986 but was associated with acute species loss. In sites fully encroached by forest, plant species richness declined by 27%, and ant richness declined by 35%. Richness of savanna specialists, the species most at risk of local extinction due to forest encroachment, declined by 67% for plants and 86% for ants. This loss highlights the important role of fire in maintaining biodiversity in tropical savannas, a role that is not reflected in current policies of fire suppression throughout the Brazilian Cerrado. In tropical grasslands and savannas throughout the tropics, carbon mitigation programs that promote forest cover cannot be assumed to provide net benefits for conservation.
–
Woody encroachment and its consequences on hydrological processes in the savannah
Eliane A. Honda & Giselda Durigan
Biological sciences, 2016
https://royalsocietypublishing.org/doi/10.1098/rstb.2015.0313
Tree density increased by an average annual rate of 6.7%, basal area at 5.7% and rain interception by the canopies at 0.6% of the gross rainfall. Independent of the vegetation structure, we found a robust relationship of 0.9% less rainfall reaching the ground as TBA increases by 1 m2 ha−1. Increases in tree biomass with woody encroachment may potentially result in less water available for uptake by plants and to recharge rivers and groundwater reserves. Given that water is a seasonally scarce resource in all savannahs, woody encroachment may threaten the ecosystem services related to water resources.
–
Europe’s forest management did not mitigate climate warming
Naudts et al.
Science, 2016
https://science.sciencemag.org/content/351/6273/597
Afforestation and forest management are considered to be key instruments in mitigating climate change. Here we show that since 1750, in spite of considerable afforestation, wood extraction has led to Europe’s forests accumulating a carbon debt of 3.1 petagrams of carbon. We found that afforestation is responsible for an increase of 0.12 watts per square meter in the radiative imbalance at the top of the atmosphere, whereas an increase of 0.12 kelvin in summertime atmospheric boundary layer temperature was mainly caused by species conversion. Thus, two and a half centuries of forest management in Europe have not cooled the climate. The political imperative to mitigate climate change through afforestation and forest management therefore risks failure, unless it is recognized that not all forestry contributes to climate change mitigation.
–
Afforestation in China cools local land surface temperature
Peng et al.
PNAS, 2014
https://www.pnas.org/content/111/8/2915
China has the largest afforested area in the world. Afforestation not only contributes to increased carbon storage but also alters local albedo and turbulent energy fluxes, which offers feedback on the local and regional climate. This study presents previously unidentified observational evidence of the effect of large-scale afforestation on land surface temperature (LST) in China. Afforestation decreases daytime LST, because of enhanced evapotranspiration, and increases nighttime LST. This nighttime warming tends to offset daytime cooling in dry regions. These results suggest it is necessary to carefully consider where to plant trees to achieve potential climatic benefits in future afforestation projects.
–
Reforester ou densifier des zones forestières peut avoir des conséquences pas forcément bénéfiques.
Cascading biodiversity and functional consequences of a global change–induced biome switch
Parr et al.
Biodiviersity research, 2012
The potential loss of many specialist savanna plant species is especially concerning, given the spatial extent and speed of this vegetation switch. Although it is not clear how easily the habitat switch can be reversed and how stable the thicket habitats are, it is likely in the not‐too‐distant future that conservation managers will be forced to make decisions on whether to actively maintain savannas.
–
Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools
Seto et al.
PNAS, 2012
https://www.pnas.org/content/109/40/16083
If current trends in population density continue and all areas with high probabilities of urban expansion undergo change, then by 2030, urban land cover will increase by 1.2 million km2, nearly tripling the global urban land area circa 2000. This increase would result in considerable loss of habitats in key biodiversity hotspots, with the highest rates of forecasted urban growth to take place in regions that were relatively undisturbed by urban development in 2000: the Eastern Afromontane, the Guinean Forests of West Africa, and the Western Ghats and Sri Lanka hotspots. Within the pan-tropics, loss in vegetation biomass from areas with high probability of urban expansion is estimated to be 1.38 PgC (0.05 PgC yr−1), equal to ∼5% of emissions from tropical deforestation and land-use change.
–
Bois et déboisement dans la Méditerranée antique
William V. Harris
Editions de l’EHESS, 2011
https://www.cairn.info/revue-annales-2011-1-page-105.htm
L’objectif premier de cet article, qui relève d’une recherche en cours, est de déterminer si la Méditerranée antique a été l’objet de déboisements à grande échelle entre 800 av. J.-C. (la naissance de la Cité grecque) et le VIIe siècle (les invasions arabes) – ce qui demeure « la question la plus controversée de l’histoire environnementale de la Méditerranée. Le second objectif est de voir s’il est possible d’écrire une histoire environnementale qui ait du sens pour une période qui, bien que brève à l’échelle des temps géologique et paléontologique, est suffisamment longue pour avoir subi des changements anthropiques majeurs.
–
The prehistoric and preindustrial deforestation of Europe
Kaplan et al.
Quaternary science review, 2009
https://www.sciencedirect.com/science/article/abs/pii/S027737910900331X
Humans have transformed Europe’s landscapes since the establishment of the first agricultural societies in the mid-Holocene. The most important anthropogenic alteration of the natural environment was the clearing of forests to establish cropland and pasture, and the exploitation of forests for fuel wood and construction materials.
–
Precipitation on land versus distance from the ocean: Evidence for a forest pump of atmospheric moisture
Makarieva et al.
Ecological complexity, 2009
https://www.sciencedirect.com/science/article/pii/S1476945X08000834
Our results indicate that forest cover plays a major role in the atmospheric circulation and water cycling on land. This suggests a good potential for forest-mediated solutions of the global desertification and water security problems.
–
Why Large-Scale Afforestation Efforts in China Have Failed To Solve the Desertification Problem
Shixiong Cao
Environmental science and technology, 2008
https://www.researchgate.net/publication/5442299_Why_Large-Scale_Afforestation_Efforts_in_China_Have_Failed_To_Solve_the_Desertification_Problem
Traditional Chinese approaches to ecosystem restoration have focused on afforestation as an important tool for controlling desertification. However, the long-term results of this practice increasingly show that these projects are actually increasing environmental degradation in arid and semiarid regions, with ecosystems deteriorating and wind erosion increasing. Rather than focusing solely on afforestation, it would be more effective to focus on recreating natural ecosystems that are more suitable for local environments and that can thus provide a better chance of combating desertification. Arid and semiarid regions make up ~40% of the earth’s land surface and are home to ~20% of the human population, but these areas are increasingly being affected by desertification (1). A half-century policy of forest exploitation, livestock overgraz-Trenches were dug parallel to the contours during planting to prevent downslope erosion and collect slope runoff for the trees. The decreased vegetation cover can offset this advantage by increasing wind erosion.
–
Planted Forests and Biodiversity
Carnus et al.
Journal of forestry, 2006
https://academic.oup.com/jof/article/104/2/65/4599206
Managing plantations to produce goods such as timber while also enhancing ecological services such as biodiversity involves tradeoffs, which can be made only with a clear understanding of the ecological context of plantations in the broader landscape and agreement among stakeholders on the desired balance of goods and ecological services from plantations.
–
Trading Water for Carbon with Biological Carbon Sequestration
Jackson et al.
Science, 2005
https://science.sciencemag.org/content/310/5756/1944
Carbon sequestration strategies highlight tree plantations without considering their full environmental consequences. We combined field research, synthesis of more than 600 observations, and climate and economic modeling to document substantial losses in stream flow, and increased soil salinization and acidification, with afforestation. Plantations decreased stream flow by 227 millimeters per year globally (52%), with 13% of streams drying completely for at least 1 year. Regional modeling of U.S. plantation scenarios suggests that climate feedbacks are unlikely to offset such water losses and could exacerbate them. Plantations can help control groundwater recharge and upwelling but reduce stream flow and salinize and acidify some soils.
–
Change in soil carbon following afforestation
Paul et al.
Forest ecology & management, 2002
https://www.sciencedirect.com/science/article/abs/pii/S037811270100740X
The most important factors affecting change in soil C were previous land use, climate and the type of forest established. Results suggest that most soil C was lost when softwoods, particularly Pinus radiata plantations, were established on ex-improved pastoral land in temperate regions. Accumulation of soil C was greatest when deciduous hardwoods, or N2-fixing species (either as an understorey or as a plantation), were established on ex-cropped land in tropical or subtropical regions. Long-term management regimes (e.g. stocking, weed control, thinning, fertiliser application and fire management) may also influence accumulation of soil C. Accumulation is maximised by maintaining longer (20–50 years) forest rotations. Furthermore, inclusion of litter in calculations reversed the observed average decrease in soil C, so that amount of C in soil and litter layer was greater than under preceding pasture.
–
Conifer Plantations on Drained Peatlands in Britain: a Net Gain or Loss of Carbon?
Cannell et al.
Forestry, 1993
https://academic.oup.com/forestry/article-abstract/66/4/353/568342?redirectedFrom=fulltext
Scenarios are presented for the time course of CO 2 -carbon gain and loss when peatlands are drained and planted with conifers. If CO 2 loss rates from drained peats are 50–100 g C m −2 a −1 there is likely to be increased carbon storage in the whole system for at least three rotations; but if CO 2 loss rates are 200–300 g C m −2 a −1 increased storage may be restricted to the first rotation, after which there is a net loss of carbon.
–
Biodiversité et teritoires agricoles
Jean-Claude Lefeuvre
Economie rurale, 1992
https://www.persee.fr/doc/ecoru_0013-0559_1992_num_208_1_4458
dans certains cas, les paysages et les structures agraires conduisent au contraire à un renforcement de la biodiversité.
Divers
The megabiota are disproportionately important for biosphere functioning
Enquist et al.
Nature communications, 2020
https://www.nature.com/articles/s41467-020-14369-y
Here, we extend metabolic scaling theory and use global simulation models to demonstrate that (i) megabiota are more prone to extinction due to human land use, hunting, and climate change; (ii) loss of megabiota has a negative impact on ecosystem metabolism and functioning; and (iii) their reduction has and will continue to significantly decrease biosphere functioning. Global simulations show that continued loss of large animals alone could lead to a 44%, 18% and 92% reduction in terrestrial heterotrophic biomass, metabolism, and fertility respectively. Our findings suggest that policies that emphasize the promotion of large trees and animals will have disproportionate impact on biodiversity, ecosystem processes, and climate mitigation.
–
Gradual replacement of wild bees by honeybees in flowers of the Mediterranean Basin over the last 50 years
Carlos M. Herrra
Proceedings of the royal society B, 2020
https://royalsocietypublishing.org/doi/10.1098/rspb.2019.2657
The proportion of wild bees at flowers was four times greater than that of honeybees at the beginning of the period, the proportions of both groups becoming roughly similar 50 years later. The Mediterranean Basin is a world biodiversity hotspot for wild bees and wild bee-pollinated plants, and the ubiquitous rise of honeybees to dominance as pollinators could in the long run undermine the diversity of plants and wild bees in the region.
–
Trophic rewilding revives biotic resistance to shrub invasion
Guyton et al.
Nature ecology and evolution, 2020
https://www.nature.com/articles/s41559-019-1068-y
Here we show that war-induced collapse of large-mammal populations in Mozambique’s Gorongosa National Park exacerbated woody encroachment by the invasive shrub Mimosa pigra—considered one of the world’s 100 worst invasive species—and that one decade of concerted trophic rewilding restored this invasion to pre-war baseline levels. Mimosa occurrence increased between 1972 and 2015, a period encompassing the near extirpation of large herbivores during the Mozambican Civil War. From 2015 to 2019, mimosa abundance declined as ungulate biomass recovered. DNA metabarcoding revealed that ruminant herbivores fed heavily on mimosa, and experimental exclosures confirmed the causal role of mammalian herbivory in containing shrub encroachment. Our results provide mechanistic evidence that trophic rewilding has rapidly revived a key ecosystem function (biotic resistance to a notorious woody invader), underscoring the potential for restoring ecological health in degraded protected areas.
–
Increased atmospheric vapor pressure deficit reduces global vegetation growth
Yuan et al.
Science advances, 2019
https://advances.sciencemag.org/content/5/8/eaax1396
Atmospheric vapor pressure deficit (VPD) is a critical variable in determining plant photosynthesis. Synthesis of four global climate datasets reveals a sharp increase of VPD after the late 1990s. In response, the vegetation greening trend indicated by a satellite-derived vegetation index (GIMMS3g), which was evident before the late 1990s, was subsequently stalled or reversed. Terrestrial gross primary production derived from two satellite-based models (revised EC-LUE and MODIS) exhibits persistent and widespread decreases after the late 1990s due to increased VPD, which offset the positive CO2 fertilization effect. Six Earth system models have consistently projected continuous increases of VPD throughout the current century. Our results highlight that the impacts of VPD on vegetation growth should be adequately considered to assess ecosystem responses to future climate conditions.
–
Global distribution of earthworm diversity
Philipps et al.
Science, 2019
Cliquer pour accéder à N525649PP.pdf
https://www.biorxiv.org/content/biorxiv/early/2019/04/09/587394.full.pdf(pas de lien direct)

–
Wild pollinator activity negatively related to honey bee colony densities in urban context
Ropars et al.
PLOS One, 2019
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0222316
Here, we show that in the city of Paris (France), wild pollinator visitation rates are negatively correlated to honey bee colony densities present in the surrounding landscape
–
Living planet report
WWF, 2018
https://wwf.panda.org/knowledge_hub/all_publications/living_planet_report_2018/
While climate change is a growing threat, the main drivers of biodiversity decline continue to be the overexploitation of species, agriculture and land conversion. Indeed, a recent assessment found that only a quarter of land on Earth is substantively free of the impacts of human activities. This is projected to decline to just one-tenth by 2050. Land degradation includes forest loss; while globally this loss has slowed due to reforestation and plantations it has accelerated in tropical forests that contain some of the highest levels of biodiversity on Earth. Ongoing degradation has many impacts on species, the quality of habitats and the functioning of ecosystems.
–
Global drivers of population density in terrestrial vertebrates
Santini et al.
Global ecology and biogeography, 2018
Cliquer pour accéder à Santini2018GEB.pdf
Traits determine most of the variation in population density across species,whereas environmental conditions explain the intraspecific variation across populations. Speciestraits, resource availability and climatic stability have a different influence on the population den-sity of the four groups. These models can be used to predict the average species populationdensity over large areas and be used to explore macroecological patterns and inform conservationanalyses.
–
What conservation does
Laurent Godet & Vincent Devictor
Trends in ecology & evolution, 2018
https://www.cell.com/trends/ecology-evolution/fulltext/S0169-5347(18)30164-2
But our empirical approach provides sufficient arguments to conclude that conservation biology does not accumulate studies dedicated to strict nature protection with no or little connexion to human well-being. Rather, we show that
several targets including the protection of nature for itself, nature for people, and nature with people are coexisting [8]. Interestingly, among most effective conservation actions, protected areas are still playing a major positive role. More generally, as the world becomes ever more dominated by humans, conservation of species in the wild is in fact even more relevant.
[…]We thus concur with adopting more lucid approaches to existing and nonreducible conflicts between biodiversity conservation and the deny of any limits to destructive human activities. Calling for new directions to conservation is neither necessary nor sufficient. Constantly seeking even more compromise when nature protection is at stake is part of the major obstacles, not of the solutions.
–
The Diversity-Weighted Living Planet Index: Controlling for Taxonomic Bias in a Global Biodiversity Indicator
McRae et al.
PLOS One, 2017
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0169156
Based on a proportionally weighted index, we estimate a global population decline in vertebrate species between 1970 and 2012 of 58% rather than 20% from an index with no proportional weighting. From this data set, comprising 14,152 populations of 3,706 species from 3,095 data sources, we also find that freshwater populations have declined by 81%, marine populations by 36%, and terrestrial populations by 38% when using proportional weighting (compared to trends of -46%, +12% and +15% respectively). These results not only show starker declines than previously estimated, but suggests that those species for which there is poorer data coverage may be declining more rapidly.
–
Humans as a Hyperkeystone Species
WOrm et al.
Trends in ecology and evolution, 2016
https://www.sciencedirect.com/science/article/pii/S0169534716300659
Ecologists have identified numerous keystone species, defined as organisms that have outsized ecological impacts relative to their biomass. Here we identify human beings as a higher-order or ‘hyperkeystone’ species that drives complex interaction chains by affecting other keystone actors across different habitats. Strong indirect effects and a global reach further characterize these interactions and amplify the impacts of human activities on diverse ecosystems, from oceans to forests. We require better understanding of hyperkeystone interaction chains most urgently, especially for marine species and terrestrial large carnivores, which experience relatively higher exploitation rates than other species. This requires innovative approaches that integrate the study of human behavior with food-web theory, and which might provide surprising new insights into the complex ecology of our own species.
–
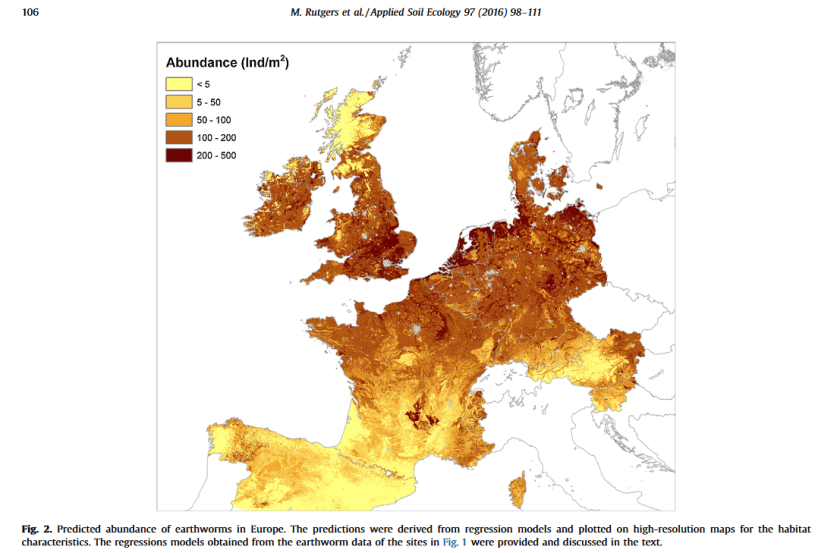
Mapping earthworm communities in Europe
Rutgers et al.
Applied soil ecology, 2016
Cliquer pour accéder à 2016.%20Rutgers.%20Mapping%20earthworm%20communities.pdf
–
A global assessment of the social and conservation outcomes of protected areas
Oldekop et al.
Conservation biology, 2015
https://conbio.onlinelibrary.wiley.com/doi/full/10.1111/cobi.12568
Whereas the strictest regimes of PA management attempted to exclude anthropogenic influences to achieve biological conservation objectives, PAs that explicitly integrated local people as stakeholders tended to be more effective at achieving joint biological conservation and socioeconomic development outcomes. Strict protection may be needed in some circumstances, yet our results demonstrate that conservation and development objectives can be synergistic and highlight management strategies that increase the probability of maximizing both conservation performance and development outcomes of PAs.
–
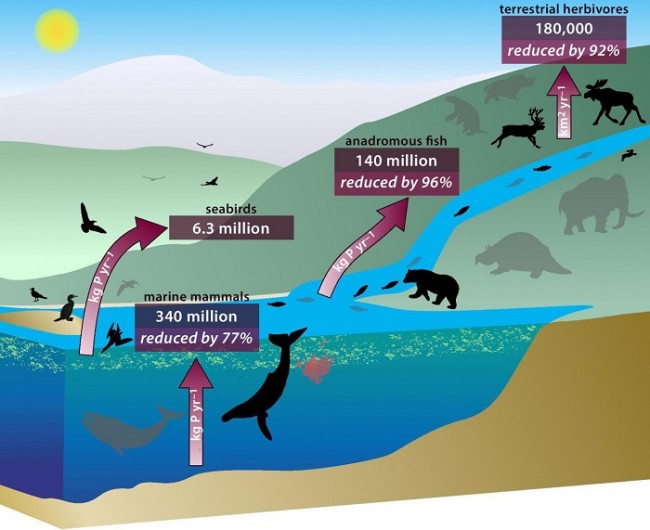
Global nutrient transport in a world of giants
Doughty et al.
PNAS 2015
Cliquer pour accéder à 1502549112.full.pdf
The past was a world of giants, with abundant whales in the sea and large animals roaming the land. However, that world came to an end following massive late-Quaternary megafauna extinctions on land and widespread population reductions in great whale populations over the past few centuries. These losses are likely to have had important consequences for broad-scale nutrient cycling, because recent literature suggests that large animals disproportionately drive nutrient movement. We estimate that the capacity of animals to move nutrients away from concentration patches has decreased to about 8% of the preextinction value on land and about 5% of historic values in oceans
–
WILDLIFE COMEBACK IN EUROPE. The recovery of selected mammal and bird species
Deinet et al.
Rewilding Europe, 2013
In this report, for the first time ever, a compre-hensive, state-of-the-art and science-based, peer-re-viewed overview of the comeback of a number of selected – often iconic – wildlife species, is described and systematically analysed.[…] Wildlife will fairly quickly bounce back if we allow it to – this report shows that. With a continued and strong legal protection, an active boosting of existing wildlife populations or by reintroductions setting up new ones, a growing nature and wildlife-based tourism offer, combined with an increasing tolerance towards wildlife, more species will surely follow. […] We find that wildlife comeback in Europe since the mid-20th century appears to be predominantly due to species protection and active targeted conservation (both birds and mammals), habitat management and site protection (birds) and legal protection (both). Of the species management techniques, actively boosting existing or setting up new populations, via translocations and reintroductions, was the foremost type of species management linked to increased abundances amongst mammals and birds. Reduction in hunting pressure, protection from persecution and the phasing out of certain toxic chemicals, thus decreasing non-natural mortality, were also important for species recovery.
–
Global meta-analysis reveals no net change in local-scale plant biodiversity over time
Vellend et al.
PNAS, 2013
https://www.pnas.org/content/110/48/19456
Finally, although plant species diversity shows no net directional change at the local scale, individual sites show tremendous variation in the direction and magnitude of biodiversity trends (Figs. 2 and 3), as well as marked changes in the identities, functional characteristics, and abundances of locally co-occurring species (i.e., species composition) (16). These changes are expected to have important consequences for ecosystem function (16, 34), and our results point to a clear need for greater attention to be paid to the consequences of local increases in diversity, which are just as common as decreases, and to changes in species composition independent of diversity changes. How often such real world changes cause ecosystem function to deteriorate, or alternatively to actually improve, remains unknown. Through continued and improved biodiversity monitoring, and experiments that simulate real world changes at the local scale, a more comprehensive understanding of biodiversity change and its consequences can be achieved.
–
The crop yield gap between organic and conventional agriculture
De Ponti et al.
Agricultural systems, 2012
https://www.sciencedirect.com/science/article/pii/S0308521X1100182X
Our review and meta-analysis of yield data comparing organic and conventional agriculture showed that currently organic yields of individual crops are on average 80% of conventional yields. The analysis of 362 datasets also showed a high variation of the yield gap of organic agriculture (standard deviation 21%). Some of this variation seems systematic. Relative yields differed between crops with e.g. soybean, some other pulses, rice and corn scoring higher than 80% and wheat, barley and potato scoring lower than 80%. Most regions have relative yields fairly close to the overall average, but Asia and Central Europe had comparatively higher and Northern Europe lower relative yields. In Denmark and The Netherlands, countries with very intensive agricultural systems, the organic–conventional yield gap was somewhat larger.
–
The Anthropogenic Greenhouse Era Began Thousands of Years Ago
William F. Ruddiman
Climatic change, 2003
https://link.springer.com/article/10.1023%2FB%3ACLIM.0000004577.17928.fa
A wide array of archeological, cultural, historical and geologic evidence points to viable explanations tied to anthropogenic changes resulting from early agriculture in Eurasia, including the start of forest clearance by 8000 years ago and of rice irrigation by 5000 years ago. In recent millennia, the estimated warming caused by these early gas emissions reached a global-mean value of ∼ 0.8 °C and roughly 2 °C at high latitudes, large enough to have stopped a glaciation of northeastern Canada predicted by two kinds of climatic models.
–
