
1. Généralités
2. Neandertal [Page dédiée]
3. Sapiens au paléolithique
4. Chasseurs-cueilleurs récents
5. Depuis le néolithique [Page dédiée]
Généralités
Cooking shapes the structure and function of the gut microbiome [Texte]
Carmody et al.
Nature, 2019
Because cooking is human-specific, ubiquitous and ancient6,7, our results prompt the hypothesis that humans and our microbiomes co-evolved under unique cooking-related pressures.
–
How carnivorous are we? The implication for protein consumption [PDF]
Miki Ben-Dor
Journal of evolution and health, 2019
A drastic decline in terrestrial mammals took place from approximately 500 kgs at the beginning of the Pleistocene 2.5 million years ago to about 10 kgs today. […]
Although we are undoubtedly omnivores, the biologic evidence that was presented here claims to show that we evolved, quite early in our evolution as the genus Homo, to become highly carnivorous and that we continue to retain a biologic adaptation to carnivory. This high level of carnivory means that during a large part of our evolution our diet was high in protein besides being high in fat.
–
Putrid Meat and Fish in the Eurasian Middle and Upper Paleolithic : Are We Missing a Key Part of Neanderthal and Modern Human Diet? [PDF]
John D. Speth
PaleoAnthropology, 2017
If such food practices were in fact widespread during the mid- to late Pleistocene, they may help account for aspects of the archaeological record that are presently difficult to comprehend, such as the ‘on again, off again’ evidence for fire use (and hence cooking) during the Eurasian Middle Paleolithic. Putrefaction also may alter the isotopic composition of the diet. As meat and fish decompose, a variety of volatile compounds are produced, including ammonia. Loss of NH3, along with lesser amounts of two other nitrogenous gases—cadaverine and putrescine—would very likely leave rotted meat and fish enriched in 15N by comparison to the isotopic composition of these foods in their fresh state. Such enrichment may have contributed to the elevated values seen in many Neanderthals, values that are widely taken as prima facie evidence of Neanderthal’s status as a ‘top predator.’ Finally, if Paleolithic foragers relied upon putrefaction to prepare and store meat, archaeologists may have to rethink the way they interpret a number of widely used taphonomic signatures, including the number and distribution of cutmarks, the extent of carnivore damage, the incidence of burning on both animal bones and stone tools, and the frequency and scale of hearths, ash lenses, and other features of combustion.
–
Evidence for chronic omega-3 fatty acids and ascorbic acid deficiency in Palaeolithic hominins in Europe at the emergence of cannibalism [PDF]
J.L. Guil-Guerrero
Quaternary science reviews, 2017
hominins at the M/UP transition had a deficit of both omega-3 fatty acids and ascorbic acid. Data on human organs summarized here are also conclusive: these contain such nutrients in amounts much higher than reached in the corresponding mammal organs consumed, and thus could have been alternative sources of those nutrients for Palaeolithic hominins. Therefore, nutritional cannibalism detected at such times could have had the function of alleviating these deficits. The evolutionary advantages gained by the consumption of the various omega-3 fatty acids of human origin are also discussed.
–
The origins and significance of coastal resource use in Africa and Western Eurasia [PDF]
Curtis Marean
Journal of human evolution, 2014
This paper provides a critical review of where and when systematic use of coastal resources and coastal adaptations appeared in the Old World by a comparison of the records from Africa and Europe. It is found that during the Middle Stone Age in South Africa there is evidence that true coastal adaptations developed while there is, so far, a lack of evidence for even the lowest levels of systematic coastal resource use by Neanderthals in Europe.
–
Hormones, 2003
When applied to the Palaeolithic-Neolithic transition, the isotopes broadly show a diet rich in protein at the end of the glacial period, a diversification, with emphasis on fish resources during the Mesolithic, and an impoverishment during the Neolithic.[…] the amount of meat eaten decreased to about 10-20% of the Upper Palaeolithic optimum.
–
Dietary lean red meat and human evolution [PDF]
Neil Mann
European journal of nutrition, 2000
In our own studies, we have shown evidence that diets high in lean red meat can actually lower plasma cholesterol, contribute significantly to tissue omega-3 fatty acid and provide a good source of iron, zinc and vitamin B12. A study of human and pre-human diet history shows that for a period of at least 2 million years the human ancestral line had been consuming increasing quantities of meat. During that time, evolutionary selection was in action, adapting our genetic make up and hence our physiological features to a diet high in lean meat. This meat was wild game meat, low in total and saturated fat and relatively rich in polyunsaturated fatty acids (PUFA).
Néandertal
Sapiens au paléolithique
Isotopic evidence of high reliance on plant food among Later Stone Age hunter-gatherers at Taforalt, Morocco [Texte]
Moubtahij et al. (Jaouen)
Nature Ecology and Evolution, 2024
–
Trophic ecology of a Late Pleistocene early modern human from tropical Southeast Asia inferred from zinc isotopes [Texte]
Bourgon et al. (Jaouen)
Journal of human evolution, 2021
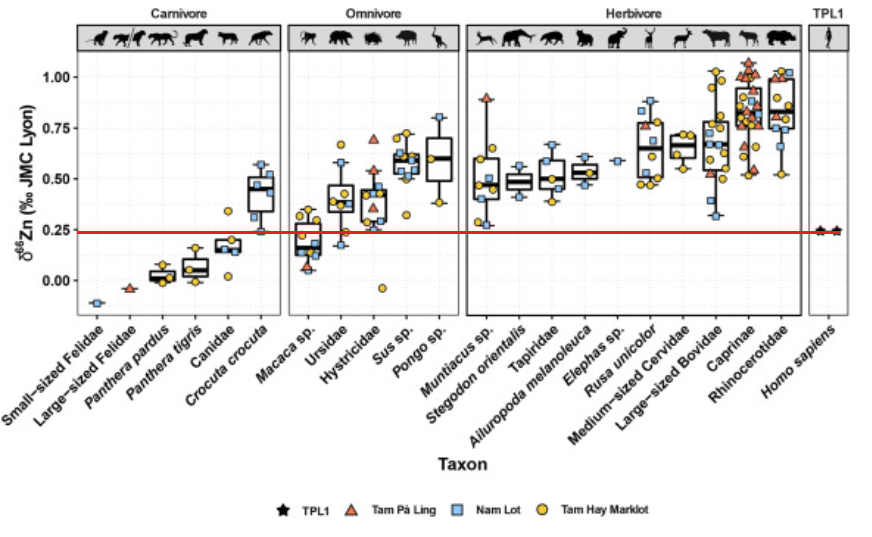
–
Stable isotopes reveal patterns of diet and mobility in the last Neandertals and first modern humans in Europe [Texte]
Wißing et al.
Nature Scientific Reports, 2019
Using the stable isotopic approach, we shed light on aspects of diet and mobility of the late Neandertals and UPMHs from the cave sites of the Troisième caverne of Goyet and Spy in Belgium. We demonstrate that their diet was essentially similar, relying on the same terrestrial herbivores, whereas mobility strategies indicate considerable differences between Neandertal groups, as well as in comparison to UPMHs. Our results indicate that UPMHs exploited their environment to a greater extent than Neandertals and support the hypothesis that UPMHs had a substantial impact not only on the population dynamics of large mammals but also on the whole structure of the ecosystem since their initial arrival in Europe.
–
The earliest direct evidence of mammoth hunting in Central Europe – The Kraków Spadzista site (Poland) [Texte]
Wojtal et al.
Quaternary science review, 2019
The oldest unequivocal evidence of mammoth hunting in prehistoric Central Europe has been found in the Gravettian archaeological site Kraków Spadzista (Poland). The site contains thousands of lithic artifacts and the remains of >100 woolly mammoths (Mammuthus primigenius), with radiocarbon dates clustering ∼25–24 ka uncal BP. A fragment of a flint shouldered point is embedded in a mammoth rib, and more than 50% of the site’s flint shouldered points and backed blades bear diagnostic traces of hafting and impact damage from use as spear tips. Additional support for mammoth killing is the mortality profile of 112 mammoths from the site: some age groups may have been depleted due to recurring heavy hunting by humans during periods of environmental stress. The evidence for intensive human hunting could portend a development thousands of years later, at the end of the Pleistocene, when climate-caused habitat changes were more extreme, and, in combination with opportunistic human hunting, may have led to woolly mammoth extinction.
–
Elephant and Mammoth Hunting during the Paleolithic: A Review of the Relevant Archaeological, Ethnographic and Ethno-Historical Records [PDF]
Agam & Barkai
Quaternary, 2018
This study examines the archaeological evidence of proboscidean hunting during Paleolithic times, and provides a review of ethnographic and ethno-historical accounts, demonstrating a wide range of traditional elephant-hunting strategies. We also discuss the rituals accompanying elephant hunting among contemporary hunter-gatherers, further stressing the importance of elephants among hunter-gatherers. Based on the gathered data, we suggest that early humans possessed the necessary abilities to actively and regularly hunt proboscideans; and performed this unique and challenging task at will.
–
Cutmark data and their implications for the planning depth of Late Pleistocene societies [Texte]
Soulier & Morin
Journal of human evolution, 2016
The present study shows that long longitudinal cutmarks are considerably more frequent during the Late Glacial Maximum than in the early Upper Paleolithic. Although the number of studies is small, actualistic data generated in controlled settings indicate that long longitudinal cutmarks are commonly produced during filleting, an activity closely associated with meat preservation, as is the case with drying and smoking. Because they provide information on possible changes in the capacity for anticipation, these results have potentially important implications for the logistical and economic organization of Paleolithic hominins.
–
Reconstruction of the Gravettian food-web at Předmostí I using multi-isotopic tracking (13C, 15N, 34S) of bone collagen [PDF]
Bocherens et al., 2015
Strong reliance on mammoth meat was found for the human of the site, similarly to previously analyzed individuals from other Gravettian sites in Moravia. Interestingly, the large canids interpreted as “Palaeolithic dogs” had a high proportion of reindeer/muskox in their diet, while consumption of mammoth would be expected from the availability of this prey especially in case of close interaction with humans. The peculiar isotopic composition of the Palaeolithic dogs of Předmostí I may indicate some control of their dietary intake by Gravettian people, who could have use them more for transportation than hunting purpose.
–
Hunters of the giants: Woolly mammoth hunting during the Gravettian in Central Europe [Extraits]
Wojtal & Wilczynsky
Quaternary international, 2015
Between 30,000 and 20,000 years ago, Gravettian hunter-gatherers spread across most of Europe. In Central Europe, large and important sites have been discovered, especially those in the Czech Republic at the base of the Pavlovské (Palava) Hills, and in southern Poland. The remains of different mammalian carnivores and herbivores accumulated in bone assemblages at these Gravettian sites. Mammoth bones and teeth are significant components in them. Mammoths certainly played a significant role in the lifetime of the Central European societies of Gravettian hunter-gatherers. These Pleistocene giants provided not only food, but also raw materials for tools and the production of ornaments. The presence of the remains of many mammoths shows that the Gravettian people were specialized in the hunting of these animals.
How do you kill 86 mammoths? Taphonomic investigations of mammoth megasites [PDF]
Pat Shipman, 2014
« The large number of individual mammoths and the scarcity of carnivore toothmarks and gnawing suggest a new ability to retain kill mammoths and control of carcasses. Age profiles of such mammoth-dominated sites with a large minimum number of individuals differ statistically at the p < 0.01 level from age profiles of Loxodonta africana populations that died of either attritional or catastrophic causes. However, age profiles from some mammoth sites exhibit a chain of linked resemblances with each other through time and space, suggesting the transmission of behavioral or technological innovation. I hypothesize that this innovation may have been facilitated by an early attempted domestication of dogs, as indicated by a group of genetically and morphologically distinct large canids which first appear in archaeological sites at about 32 ka B.P. »
–
Estimated macronutrient and fatty acid intakes from an East African Paleolithic diet [Texte]
Kuipers et al.
British Journal of Nutrition, 2010
We found (range of medians in en%) intakes of moderate-to-high protein (25–29), moderate-to-high fat (30–39) and moderate carbohydrates (39–40). The fatty acid composition was SFA (11·4–12·0), MUFA (5·6–18·5) and PUFA (8·6–15·2). The latter was high in α-linolenic acid (ALA) (3·7–4·7 en%), low in LA (2·3–3·6 en%), and high in long-chain PUFA (LCP; 4·75–25·8 g/d), LCP n-3 (2·26–17·0 g/d), LCP n-6 (2·54–8·84 g/d), ALA/LA ratio (1·12–1·64 g/g) and LCP n-3/LCP n-6 ratio (0·84–1·92 g/g).
–
Isotopic evidence for the diets of European Neanderthals and early modern humans [Texte]
Richards & Trinkaus
PNAS, 2009
Isotopic methods indicate the sources of dietary protein over many years of life, and show that Neanderthals had a similar diet through time≈120,000 to ≈37,000 cal BP) and in different regions of Europe. The isotopic evidence indicates that in all cases Neanderthals were top-level carnivores and obtained all, or most, of their dietary protein from large herbivores. In contrast, early modern humans (≈40,000 to ≈27,000 cal BP) exhibited a wider range of isotopic values, and a number of individuals had evidence for the consumption of aquatic (marine and freshwater) resources.
–
Stable isotope dietary analysis of the Tianyuan 1 early modern human [Texte]
Hu et al.
PNAS, 2009
This analysis provides the direct evidence for the consumption of aquatic resources by early modern humans in China and has implications for early modern human subsistence and demography.
Chasseurs-cueilleurs récents
Current views on hunter‐gatherer nutrition and the evolution of the human diet [Texte]
Crittenden & Schnorr
American journal of biological anthropology, 2017
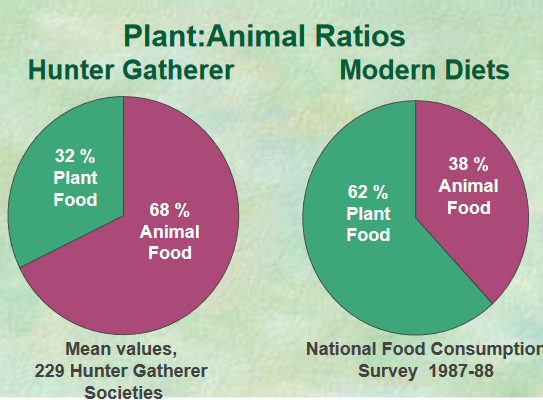
These sources suggest that plants constitute a much higher contribution to the diet, approximately 65%, with animal products making up the remaining 35% (Eaton, Shostak, & Konner, 1988). In addition, a more recent analysis by Frank Marlowe (Marlowe, 2005; and also summarized in relation to the Hadza of Tanzania in his 2010 book, The Hadza) suggests that the median diet of warm‐climate foragers is composed of 53% gathered plant foods, 26% hunted foods, and 21% fished foods.[…]
In addition, meat consumed by foraging populations is not limited to large game muscle tissue and extends to all edible portions of the carcass—inclusive of organs, bone marrow, and sometimes even the contents of the gastrointestinal tract of the animal […] Furthermore, many populations also consume variable quantities of insects, which until quite recently were often discarded or underestimated.
Diets of modern hunter-gatherers vary substantially in their carbohydrate content depending on ecoenvironments: results from an ethnographic analysis [PDF]
Ströhle & Hahn
Nutrition research, 2011
using data of plant-to-animal subsistence ratios, we calculated the carbohydrate intake (percentage of the total energy) in 229 hunter-gatherer diets throughout the world and determined how differences in ecological environments altered carbohydrate intake. We found a wide range of carbohydrate intake (≈3%-50% of the total energy intake; median and mode, 16%-22% of the total energy). Hunter-gatherer diets were characterized by an identical carbohydrate intake (30%-35% of the total energy) over a wide range of latitude intervals (11°-40° north or south of the equator). However, with increasing latitude intervals from 41° to greater than 60°, carbohydrate intake decreased markedly from approximately equal to 20% to 9% or less of the total energy. Hunter-gatherers living in desert and tropical grasslands consumed the most carbohydrates (≈29%-34% of the total energy). Diets of hunter-gatherers living in northern areas (tundra and northern coniferous forest) contained a very low carbohydrate content (≤15% of the total energy). In conclusion, diets of hunter-gatherers showed substantial variation in their carbohydrate content. Independent of the local environment, however, the range of energy intake from carbohydrates in the diets of most hunter-gatherer societies was markedly different (lower) from the amounts currently recommended for healthy humans.
–
Diet and the evolution of human amylase gene copy number variation [Texte]
Perry et al.
Nature Genetics, 2007
We found that salivary amylase gene (AMY1) copy number is correlated positively with salivary amylase protein levels, and that individuals from populations with high-starch diets have on average more AMY1 copies than those with traditionally low-starch diets.
–
Hunting and Gathering. The Human Sexual Division of Foraging Labor [Abstract]
Frank W. Marlowe
Cross-cultural research, 2007
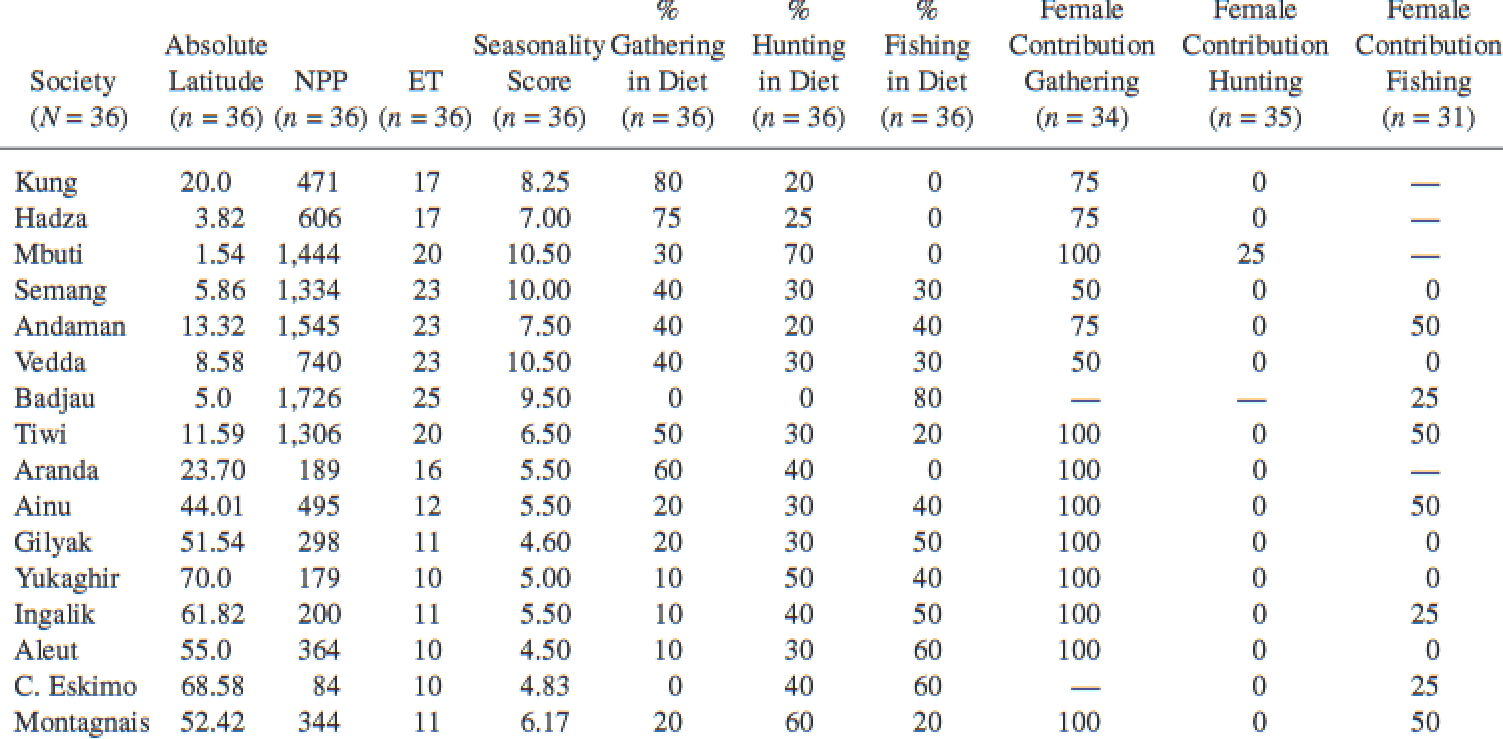
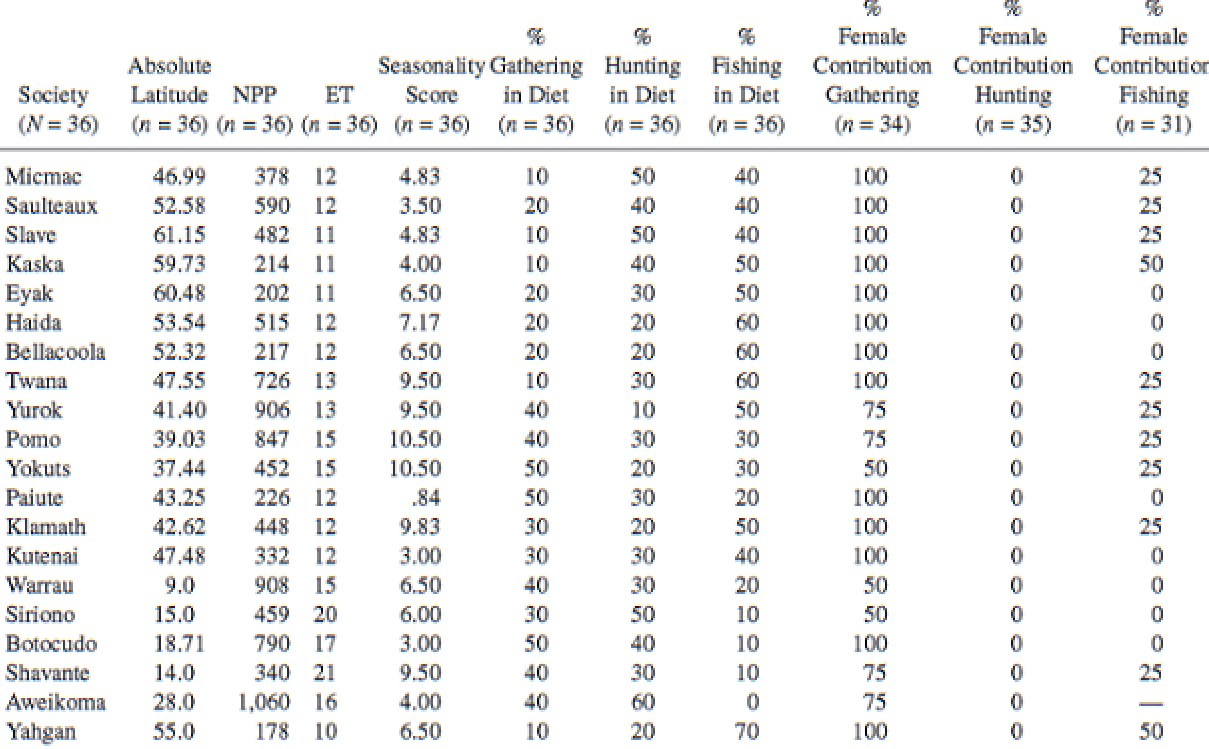
–
Hunter-Gatherers and Human Evolution [PDF]
Frank w. Marlowe
Evolutionary anthropology, 2005

Paleolithic nutrition, what did our ancestors eat ? [PDF]
Brand Miller et al, 2009
–
Evolution : Diet, Intelligence, and Longevity [PDF] [PDF]
Kaplan et al.
Evolutionnary anthropology, 2000

Plant-animal subsistence ratios and macronutrient energy estimations in worldwide hunter-gatherer diets [Texte]
Cordain et al.
The American Journal of Clinical Nutrition, 2000
Our analysis showed that whenever and wherever it was ecologically possible, hunter-gatherers consumed high amounts (45–65% of energy) of animal food. Most (73%) of the worldwide hunter-gatherer societies derived >50% (≥56–65% of energy) of their subsistence from animal foods, whereas only 14% of these societies derived >50% (≥56–65% of energy) of their subsistence from gathered plant foods. This high reliance on animal-based foods coupled with the relatively low carbohydrate content of wild plant foods produces universally characteristic macronutrient consumption ratios in which protein is elevated (19–35% of energy) at the expense of carbohydrates (22–40% of energy).
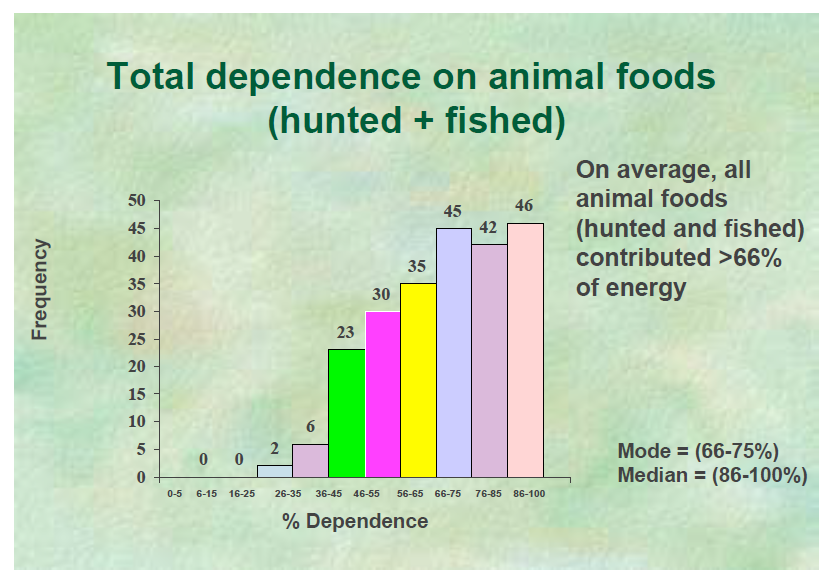
Traditional diet and food preferences of Australian Aboriginal hunter-gatherers [Abstract]

–
Paleolithic Nutrition, a consideration of its nature and current implications [PDF]
Eaton & Konner
New England journal of medicine, 1985
–
Why hunters gather: optimal foraging and the Aché of eastern Paraguay [PDF]
Hawkes et al.
American ethnologist, 1982.
–
