Il est souvent affirmé qu’Homo sapiens possède le même système digestif que les autres grands singes. Il est aussi affirmé qu’il possède une morphologie plus proche de celle des animaux herbivores que de celle des animaux carnivores. Il est par ailleurs parfois affirmé que les autres grands singes sont quasi exclusivement végétariens.
On retrouve notamment ces affirmations dans les vidéos très fortement médiatisées de Gary Yourofsky ou d’Irène Grosjean.
Qu’en est-il vraiment ?
1. Comparaison humain/autres animaux : alimentation
2. Comparaisons anatomiques
3. Comportement alimentaire des primates
4. Vulgarisation, presse
Comparaison humain/autres animaux : alimentation
Widespread extinctions of co-diversified primate gut bacterial symbionts from humans [Abstract]
Sanders et al.
Nature Mibrobiology, 2023
–
In Vitro Models of the Canine Digestive Tract as an Alternative to In Vivo Assays: Advances and Current Challenges [PDF]
Deschamps et al.
Altex, 2022
Because dogs are facultative carnivores, their GIT is adapted to high–protein and high–fat diets, i.e., relatively short and simple compared to herbivores and even omnivores like human or swin […]
Gastric pH of Beagles under fasted conditions is around 1.5 (range 0.9–2.5), quite similar to that of human (range 1.4–2.1)
–
Les humains ont évolué pour métaboliser mieux les lipides, les chimpanzés ont évolué pour métaboliser mieux les glucides :
Comparative analyses of chromatin landscape in white adipose tissue suggest humans may have less beigeing potential than other primates [Texte]
Swain-Lenz et al.
Genome biology and evolution, 2019
Taken together, these results suggest that humans shut down regions of the genome to accommodate a high fat diet while chimpanzees open regions of the genome to accommodate a high sugar diet.
–
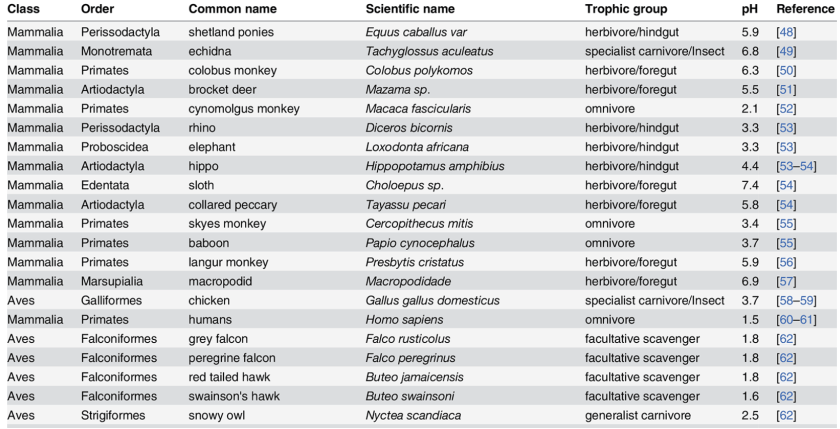
Une comparaison générale de l’acidité de l’estomac de plusieurs dizaines d’animaux, dont il ressort que l’estomac humain est en fait anormalement acide pour un omnivore, son acidité plaçant en fait Homo sapiens quelque part entre les carnivores et les charognards. Autre indice suggérant une part importante du charognage ou de la consommation de proies longtemps après capture dans l’histoire évolutive humaine :
The Evolution of Stomach Acidity and Its Relevance to the Human Microbiome [Texte]
Beasley et al
PLOS One, 2015
–
Variable responses of human and non-human primate gut microbiomes to a Western diet [Texte]
Amato et al.
Microbiome, 2015
Here, we show that the human microbiome reacts differently to a high-protein, high-fat Western diet than that of a model primate, the African green monkey, or vervet (Chlorocebus aethiops sabaeus). Specifically, humans exhibit increased relative abundance of Firmicutes and reduced relative abundance of Prevotella on a Western diet while vervets show the opposite pattern. Predictive metagenomics demonstrate an increased relative abundance of genes associated with carbohydrate metabolism in the microbiome of only humans consuming a Western diet.
–
When man met meat: meat in human nutrition from ancient times till today [PDF]
Baltic & Boskovic
Procedia food science, 2015
There is evidence that meat consumption has had an influence on cranial-dental and intestinal morphologic changes, human erect posture, reproductive characteristics, longer lifespan, and maybe most importantly, on brain and intellectual development.
–
La surface d’absorption du système digestif humain serait en réalité bien plus petite qu’on ne le pensait, ce qui ne va pas dans le sens d’une importante consommation végétale (avec la réduction globale du volume, la réduction relative très importante du côlon, et les modifications du microbiotes au cours de l’évolution humaine, voir plus bas).
Surface area of the digestive tract – revisited [Abstract]
Helander et Fändricks.
Scandinavian journal of gastroenterology, 2014
Results. Literature review of intubation or radiological methods indicates an oroanal length of ∼5 m, two-third of which refers to the small intestine. However, there is a considerable variation between individuals. The inner diameter of the small intestine averages 2.5 cm and that of the large intestine averages 4.8 cm. The mucosa of the small intestine is enlarged ∼1.6 times by the plicae circulares. Morphometric data obtained by light and electron microscopy of biopsies demonstrate that villi and microvilli together amplify the small intestinal surface area by 60–120 times. Surface amplification due to microvilli in the colon is ∼6.5 times. The mean total mucosal surface of the digestive tract interior averages ∼32 m2, of which about 2 m2 refers to the large intestine. Conclusion. The total area of the human adult gut mucosa is not in the order of tennis lawn, rather is that of half a badminton court.
–
Le microbiome des humains modernes est très différent de ce qu’il était il y a quelques millions d’années, et très différent de celui des autres primates. Il est plus adapté à une consommation de produits animaux.
Rapid changes in the gut microbiome during human evolution [Texte]
Moeller et al., 2014.
PNAS, 2014
Relative to the microbiomes of wild apes, human microbiomes have lost ancestral microbial diversity while becoming specialized for animal-based diet
–
Il est parfois affirmé que les lions ont des intestins très courts, de seulement 2m. La réalité est plutôt vers les 7-8m, proche de celle des intestins humains (mais avec un ratio longueur des intestins/longueur du corps plus faible chez le lion (ce qui semble normal pour un carnivore pur).
Aspects of Carcass Digestibility by African Lions (Panthera leo Linnaeus, 1758) under captive conditions [Texte]
Smith et al.
Pakistan Journal of Biological Sciences, 2006

–
Les premiers représentant du genre Homo ont les dents moins plates à la fois que celles des Australopithèques, mais aussi que celles des chimpanzés, ce qui serait cohérent avec une adaptation à une consommation accrue de viande.
Dental topography and diets of Australopithecus Afarensis and early Homo [Texte]
Peter Ungar
Journal of Human evolution, 2004
Early Homo shows steeper slopes and more relief than chimpanzees, whereas A. afarensis shows less slope and relief than any of the other groups. The differences between the two hominin taxa are on the same order as those between the extant apes, suggesting similar degrees of difference in diet. Because these chimpanzees and gorillas differ mostly in fallback foods where they are sympatric, results suggest that the early hominins may likewise have differed mostly in fallback foods, with A. afarensis emphasizing harder, more brittle foods, and early Homo relying on tougher, more elastic foods.
–
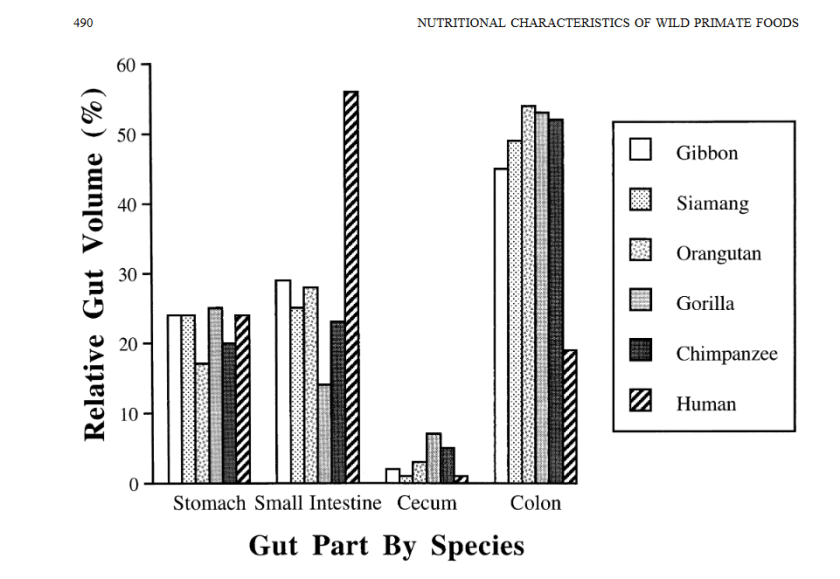
Les intestins humains diffèrent significativement des intestins des autres primates, notamment dans le volume relatif de l’intestin grêle (beaucoup plus grand chez sapiens) et dans le volume relatif du côlon (beaucoup plus petit chez sapiens), laissant entendre une adaptation à une alimentation sensiblement plus carnée. Du fait d’un volume global plus faible du système digestif dans son ensemble (voir plus bas), en réalité, le volume global du côlon humain rapporté au volume du corps entier est environ 4 fois plus faible :
Nutritional Characteristics of Wild Primate Foods: Do the Diets of Our Closest Living Relatives Have Lessons for Us ? [PDF]
Katharine Milton
Nutrition, 1999
–
Selon cette étude, il semble difficile d’affirmer que la longueur des intestins humains soit plus proche de celle des herbivores que de celle des carnivores. En réalité, elle semble assez similaire à celle du chien, et pas très éloignée de celle du chat :
Human adaptations to meat eating [Abstract]
Henneberg et al.
Human Evolution, 1998

–
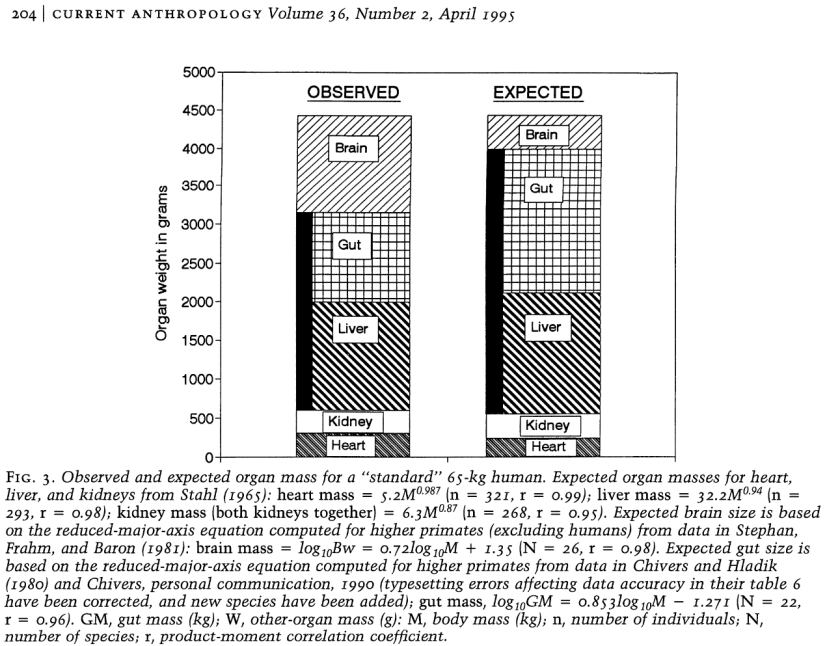
Le volume relatif des intestins humains est nettement plus faible que celui des autres primates. Cela laisse supposer une adaptation à une alimentation beaucoup plus riche.
The expensive-tissue hypothesis : the brain and the digestive system in human and primate evolution [PDF]
Aiello & Wheeler
Current anthropology, 1995
–
On sait depuis longtemps que la longueur de l’intestin n’est qu’une donnée relative, qui ne peut avoir qu’un caractère indicatif.
Des variations de longueur de l’intestin [Texte]
Adolphe Block
Bulletins et mémoires de la société d’anthropologie de Paris, 1904

Comparaisons anatomiques
Rethinking the evolution of the human foot: insights from experimental research [PDF]
Holowka & Lieberman
Journal of experimental biology, 2018
the human foot passed through three evolutionary stages: first, a great ape-like foot adapted for arboreal locomotion but with some adaptations for bipedal walking; second, a foot adapted for effective bipedal walking but retaining some arboreal grasping adaptations; and third, a human-like foot adapted for enhanced economy during long-distance walking and running that had lost its prehensility.
–
Chimpanzee super strength and human skeletal muscle evolution [PDF]
O’Neil et al.
PNAS 2017
Since at least the 1920s, it has been reported that common chim-panzees (Pantroglodytes) differ from humans in being capable of exceptional feats of “super strength” both in the wild and in captive environments. A mix of anecdotal and more controlled studies provides some support for this view; however, a critical review of available data suggests that chimpanzee mass-specific muscular performance is a more modest 1.5 times greater than humans on average […] Computer simulations of species-specific whole-muscle models indicate that maximum dynamic force and power output is 1.35 times higher in a chimpanzee muscle than a human muscle of similar size. […] . We propose that the hominin lineage experienced a decline in maximum dynamic force and power output during the past 7–8 million years in response to selection for repetitive, low-cost contractile behavior.
–
Depuis Homo erectus, les humains possèdent des membre inférieurs beaucoup plus longs et droits que ceux de leurs cousins primates. Associée à de meilleurs capacités de respiration durant la course et à de meilleures capacités de sudation et de régulation thermique, cette évolution fait d’eux de redoutables coureurs d’endurance.
Pour l’ensemble des adaptations humaines à la course d’endurance, voir cette page.
Ici, les auteurs insistent sur les évolutions qu’ils attribuent à la maîtrise du feu, y compris l’allongement des membres inférieurs, bien visible ici :
Human Adaptation to the Control of Fire [PDF]
Richard Wrangham & Rachel Carmody
Evolutionnary anthropology, 2010
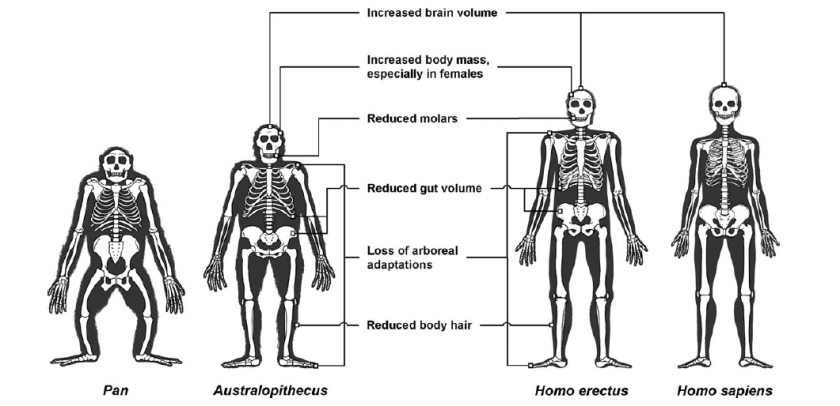
–
Unique morphology of the human eye and its adaptive meaning: comparative studies on external morphology of the primate eye [Abstract]
Kobayashi & Kohshima
Journal of human evolution, 2001
adaptations for extending the visual field by eyeball movement, especially in the horizontal direction. Comparison of eye coloration and facial coloration around the eye suggested that the dark coloration of exposed sclera of nonhuman primates is an adaptation to camouflage the gaze direction against other individuals and/or predators, and that the white sclera of the human eye is an adaptation to enhance the gaze signal. The uniqueness of human eye morphology among primates illustrates the remarkable difference between human and other primates in the ability to communicate using gaze signals.
Comportement alimentaire des primates
Meat eating by nonhuman primates: A review and synthesis [Abstract]
David P. Watts
Journal of human evolution, 2020
At least 89 nonhuman primate species representing 39 genera and 12 families eat meat. Birds are the most common prey taxon, followed by reptiles, amphibians, mammals, and fish. Most primates eat meat rarely, if at all, but meat sometimes provides substantial immediate energy and protein gains. The main importance of meat is probably as a source of vitamins, minerals, and micronutrients.
Chimpanzees eat more meat than all other nonhuman primates, but much less than African hunter-gatherers. Nonhuman primates only hunt prey much smaller than themselves. Only some capuchins and some chimpanzees use tools to assist in prey capture.
–
New Observations of Meat Eating and Sharing in Wild Bonobos (Pan paniscus) at Iyema, Lomako Forest Reserve, Democratic Republic of the Congo [Abstract]
Wakefield et al.
Folia primatologica, 2019
This report contributes to a growing body of data suggesting that wild bonobos consume meat at higher rates than previously thought, female control of carcasses is frequent but not exclusive, and meat sharing in bonobos is primarily passive but not without aggression.
–
Les macaques ne laissent pas leur part de viande aux chiens. Ils ont une prédilection pour le rat. A tel point que dans les plantations de palmiers, une étude suggère que leur effet régulateur sur les rats est supérieur à leurs nuisances sur les végétaux.
Macaques can contribute to greener practices in oil palm plantations when used as biological pest control [Texte]
Holtzner et al.
Current biology, 2019
wild macaques have the potential to act as biological pest control by feeding on plantation rats, the major pest for oil palm crops, with each macaque group estimated to reduce rat populations by about 3,000 individuals per year
–
Des chimpanzés sont capables de voler leur proie à des prédateurs de taille moyenne à grande. Cela renforce l’idée que les préhumains et premiers représentants du genre Homo étaient capables de faire de même.
Wild chimpanzees deprived a leopard of its kill: Implications for the origin of hominin confrontational scavenging [Abstract]
Nakamura et al.
Journal of human evolution, 2019
Chimpanzees do scavenge meat, although rarely, but no previous evidence of confrontational scavenging has hitherto emerged. Thus, it was assumed that they are averse to confrontation with even leopard-sized predators. However, in the observed case the chimpanzees frequently emitted waa barks, which indicated that they were aware of the leopard’s presence but they nevertheless continued to eat the scavenged meat. In addition, we compiled and reviewed 49 cases of chimpanzee encounters with animal carcasses in the Mahale Mountains of Tanzania in 1980–2017. Chimpanzees scavenged meat in 36.7% of these cases, and tended to eat the meat when it was fresh or if the animal species was usually hunted by chimpanzees. However, no evidence indicated that carcasses were avoided when leopard involvement was likely. These results suggest that chimpanzee-sized hominins could potentially confront and deprive leopard-size carnivores of meat.
–
Comparaison des pratiques de chasse des chimpanzés et des humains, et implications pour la lignée humaine depuis l’ancêtre commun avec les chimpanzés. Les humains sont à la fois plus chasseurs et plus charognards que les chimpanzés :
From Pan to Man the Hunter: Hunting and Meat Sharing by Chimpanzees, Humans, and Our Common Ancestor [PDF]
Wood & Gilby
In Chimpanzees and Human Evolution, 2018
Humans eat more meat than any other anthropoid primate, attesting to amajor shift in the diet of our hominin ancestors. Hunting and meat sharing are central to hypotheses explaining the evolution of several derived human traits, including large brains, long childhoods, small guts and teeth, complex cooperation, the sexual division of labor, cooperative breeding, and the expansion of Homo spp. around the world
–
Les chimpanzés mangent leurs jeunes proies en commençant par la cervelle, qui est la partie nutritionnellement la plus prisée.
Meat Eating by Wild Chimpanzees (Pan troglodytes schweinfurthii): Effects of Prey Age on Carcass Consumption Sequence [Texte]
Gilby & Wawrzyniak
International journal of primatology, 2018
Overall, the head was significantly more likely to be targeted first than either the torso (including viscera) or appendages. This result was driven by subadult prey, 91% of which were eaten head-first, probably because their skulls were relatively easy for chimpanzees to break with a single bite.
–
Chimpanzee vertebrate consumption: Savanna and forest chimpanzees compared [Texte]
Moore et al.
Journal of Human Evolution, 2017
There is broad consensus among paleoanthropologists that meat-eating played a key role in the evolution of Homo, but the details of where, when, and why are hotly debated. It has been argued that increased faunivory was causally connected with hominin adaptation to open, savanna habitats. […] Combining these with published fecal analyses, we summarize chimpanzee vertebrate consumption rates, showing quantitatively that savanna chimpanzee populations do not differ significantly from forest populations. Compared with forest populations, savanna chimpanzees consume smaller vertebrates that are less likely to be shared, and they do so more seasonally. […] These findings bear on discussions of why chimpanzees hunt and suggest that increases in hominin faunivory were related to differences between hominins and chimpanzees and/or differences between modern and Pliocene savanna woodland environments.
–
Mammals consumed by bonobos (Pan paniscus): new data from the Iyondji forest, Tshuapa, Democratic Republic of the Congo [PDF]
Sakamaki et al.
Primates, 2016
We found evidence that Iyondji bonobos consumed duikers (Cephalophus dorsalis,C. monticola) and diurnal monkeys (Cercopithecus asca-nius), which is notable because only anomalures (Anoma-lurusspp.) are consumed by bonobos in Wamba, a long-term study site established in 1973, located in an area adjacent to Iyondji.
–
New evidence on the tool-assisted hunting exhibited by chimpanzees (Pan troglodytes verus) in a savannah habitat at Fongoli, Sénégal [Texte]
Pruetz et al.
Royal Society Open Science, 2015
Here, we test the hypothesis that sex and age patterns in tool-assisted hunting (n=308 cases) at Fongoli occur and differ from chimpanzees elsewhere, and we compare tool-assisted hunting to the overall hunting pattern. Males accounted for 70% of all captures but hunted with tools less than expected based on their representation on hunting days. Females accounted for most tool-assisted hunting. We propose that social tolerance at Fongoli, along with the tool-assisted hunting method, permits individuals other than adult males to capture and retain control of prey, which is uncommon for chimpanzees. We assert that tool-assisted hunting could have similarly been important for early hominins.
–
Primate hunting by bonobos at LuiKotale, Salonga National Park [Texte]
Surbeck & Hohmann
Current Biology, 2008
Chimpanzees (Pan troglodytes) and bonobos (P. paniscus) hunt and consume the meat of various mammals. While chimpanzees frequently hunt in groups for arboreal, group-living monkey species, bonobos are thought to focus on medium-sized terrestrial prey, such as forest antelopes, squirrels and other rodents, which are caught opportunistically by single individuals. The absence of monkey hunting by bonobos is often used to illustrate the divergent evolution of the two Pan species. Here, we present the first information on hunting of diurnal, arboreal and group living primates by wild bonobos.
–
Selon les lieux, les chimpanzés peuvent consommer des quantités de viande importante. Les mâles en consomment nettement plus que les femelles.
Chimpanzee Hunting Behavior [PDF]
Nicholas Newton- Fischer
In Handbook of paleoanthropology, Henke & Tattersal, 2007
The 45 Gombe chimpanzees of the Kasakela community in 1992 consumed over 500 kg of red colobus meat, and their total meat consumption for the year was probably close to 700 kg. The previous year (1991), colobus meat consumption was less than 200 kg, and in 1988, this figure was less than 150 kg (Stanford 1998). Averaged over years, the level of consumption in the 1980s and 1990s seems similar to the estimate of 441 kg of meat per year for the same community in the 1970s (Wrangham and van Zinnicq Bergmann Riss 1990; Stanford 1998).Boesch and Boesch‐Achermann (2000) estimate that, averaged across the year, male Taı chimpanzees consume 186 g per day, while females consume 25 g per day. Their estimates for Gombe chimpanzees, similarly averaged, are 55 g per day for males and 7 g per day for females. These are similar to estimates made by Stanford (1998) of 70 g per day for males during peak hunting season and by Wrangham (1975) of 22 g averaged over males and females.
–
Les gorilles consomment des quantités plus ou moins importantes d’insectes.
Intrapopulation differences in ant eating in the mountain gorillas of Bwindi Impenetrable National Park, Uganda [PDF]
Ganas & Robbins
Primates, 2004
–
Selon cette étude, deux listes sont bien fournies : la liste des primates qui chassent (et qui chassent des mammifères), et la liste de leurs proies, dont font parfois partie… des enfants humains :
Le singe carnivore. La chasse chez les primates non humains [Texte]
Ducros & Ducros
Bulletins et mémoires de la société d’anthropologie de Paris, 1992


–
Insectivory by gorillas [Abstract]
Harcourt & Harcourt
Folia Primatologica, 1984
–
Chasse et usage d’outils chez les chimpanzés, déjà connus en 1981.
Hunter, gatherers, and sex roles in space and time [Texte]
Nancy Makepeace Tanner
Recension de Woman the Gatherer, Frances Dahlberg, 1981
American Anthropologist, 1983


–
L’alimentation protéique du Chimpanzé dans son environnement forestier naturel [PDF]
Hladik & Viroben
Comptes-rendus de l’Académie des Sciences de Paris, 1974
Vulgarisation, presse blogs…
L’être humain, herbivore ou carnivore ? [Page]
Quebec Science, 2018
Sur le PH, contradiction avec Beasley et al., 2015.
Toutefois, il faut savoir que l’homme n’a pas non plus un système digestif de carnivore! En effet, le pH ton estomac n’est pas aussi acide que celui des véritables mangeurs de viande.
–
L’homme est-il omnivore ou herbivore ? [Site apparemment supprimé]
Aventure-vegan, 2017
CONCLUSION
Ce qui est certain à ce stade, pour en revenir à la raison et à la mission de l’aventure végane, c’est que le fait d’adopter un régime végétarien ou végétalien au minimum, et végan au mieux, est en harmonie complète avec notre nature. En revanche, le fait de manger de la chair, que ce soit de la viande ou du poisson, est bel et bien contre nature. Voilà déjà une réponse claire obtenue à l’une de nos préoccupations.
–
Une représentation naïve d’anatomie comparée se basant sur des auteurs du 19ème siècle :