Longévité et espérance de vie sont deux notions différentes. L’espérance de vie fait référence à l’âge moyen auquel meurt un individu. Elle est généralement de 30 à 40 ans au maximum chez les chasseurs-cueilleurs, et plus généralement dans l’ensemble des sociétés précédant la révolution industrielle, et de l’ordre de 80 ans dans les sociétés industrielles. La longévité est l’âge auquel peut prétendre parvenir un individu s’il échappe aux différentes causes de mortalité accidentelles. Elle est de l’ordre de 70 à 80 ans chez les chasseurs-cueilleurs. La différence entre les deux valeurs s’explique par l’importante mortalité infantile que connaissent les sociétés pré-industrielles, et par une mortalité plus modeste, mais toujours supérieure à celle des sociétés industrielles, tout au long de l’âge adulte.
Voir aussi ce thread pour plus de détails sur cette question.
1. Longévité et espérance de vie dans les sociétés pré-industrielles.
2. Records de longévité contemporains.
3. L’hypothèse de la grand-mère.
4. Divers
Longévité et espérance de vie dans les sociétés pré-industrielles
The life history of human foraging : cross-cultural and individual variation [PDF]
Jeremy Koster et al.
Science Advances, 2020
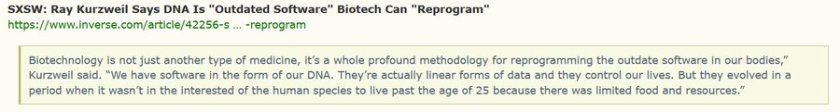
Walker et al.(2002) and Gurven et al. (2006) report data from the southern Neotropics that subsistence hunters achieve high proficiency only after reaching advanced ages, roughly 35 to 45 years old. Because hunters achieve adult size and strength much earlier in life, these results are consistent with the embodied capital hypothesis and its emphasis on the gradual mastery of cognitively complex hunting strategies […]
Our analysis supports the general conclusion that skill peaks between 30 and 35 years of age, well after the age of reproductive maturity. Peak skill is typically not much higher than skill during early adulthood, however. Declines with age are typically slow—an average 56 year old has the same proportion of maximum skill as an average 18 year old. There is considerable variation both among sites and individual hunters within study sites. Variation among individuals is described more by heterogeneity in the rate of decline than the rate of gain.
Favorable ecological circumstances promote life expectancy in chimpanzees similar to that of human hunter-gatherers [PDF]
Wood et al.
Journal of human evolution, 2017
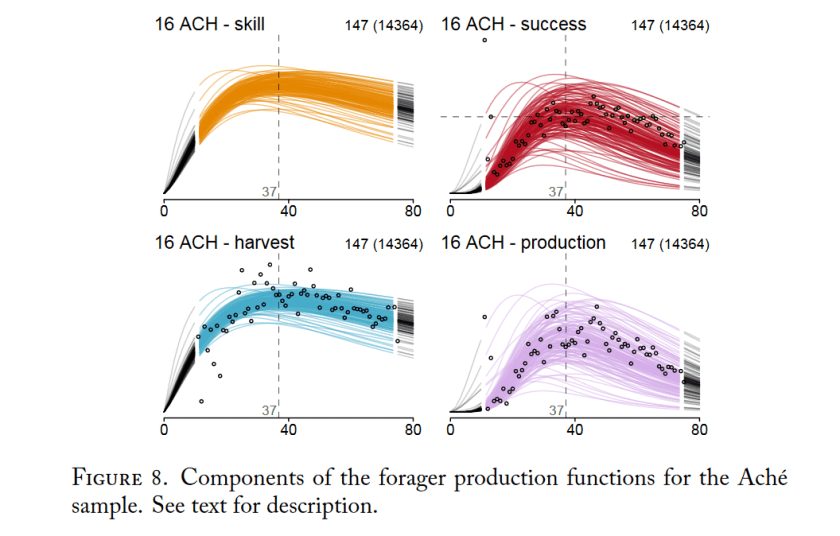
Life expectancy at birth for both sexes combined was 32.8 years, far exceeding estimates of chimpanzee life expectancy in other communities, and falling within the range of human hunter-gatherers (i.e., 27–37 years). Overall, the pattern of survivorship at Ngogo was more similar to that of human hunter-gatherers than to other chimpanzee communities. Maximum lifespan for the Ngogo chimpanzees, however, was similar to that reported at other chimpanzee research sites and was less than that of human-hunter gatherers.
Maximum likelihood estimate of life expectancy in the prehistoric Jomon: Canine pulp volume reduction suggests a longer life expectancy than previously thought [Abstract]
Sasaki & Kondo
American journal of physical anthropology, 2016
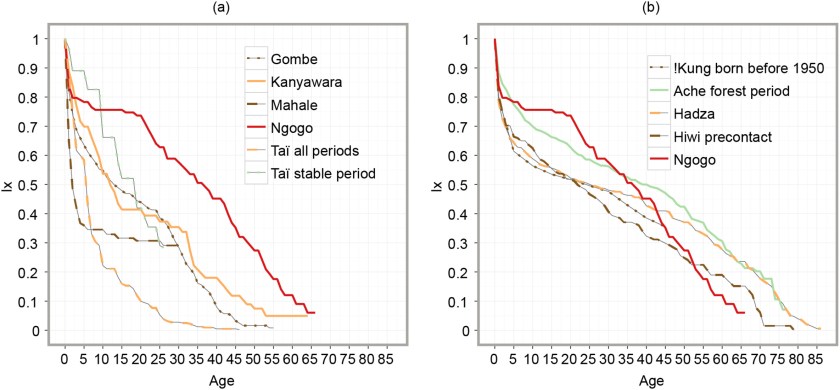
The life expectancy at age 15 of the Jomon period prehistoric populace in Japan was initially estimated to have been ∼16 years while a more recent analysis suggested 31.5 years. In this study, we provide alternative results based on a new methodology.
[…] The rate‐adjusted result of 32.2 years more likely represents the true life expectancy of the Jomon people at age 15, than the result without adjustment. Considering ∼7% rate of antemortem loss of the mandibular canine observed in our Jomon period sample, actual life expectancy at age 15 may have been as high as ∼35.3 years.
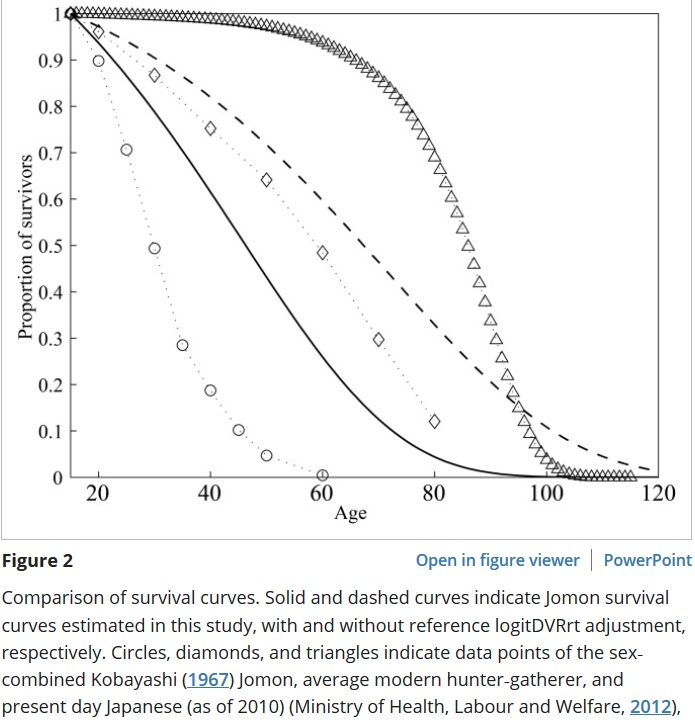
L’espérance de vie à 70 ans est encore d’une dizaine d’années chez la plupart des chasseurs-cueilleurs. Elle est encore de quelques années à 80 ans, ce qui signifie que cet âge peut même être dépassé.
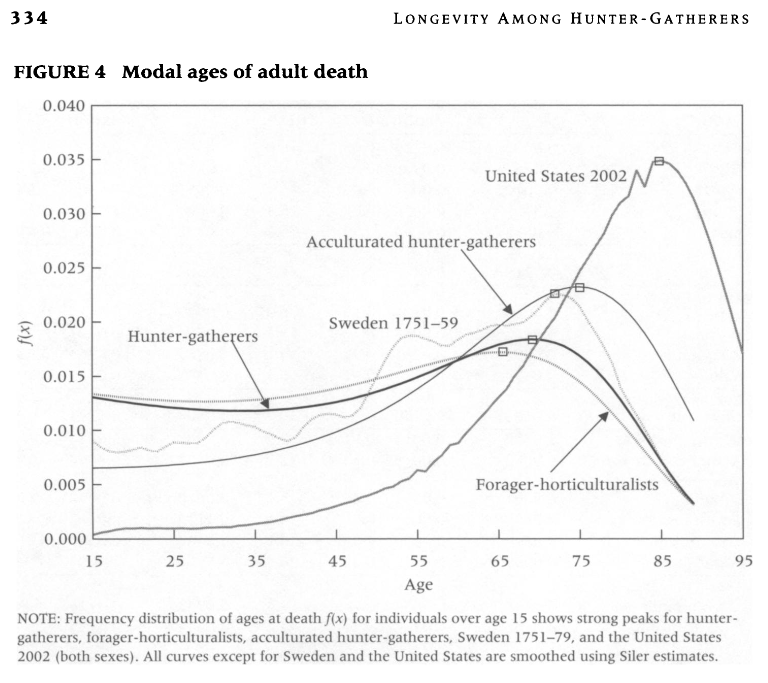
Le second graphique représente l’évolution des décès des adultes par âge. L’âge modal au décès est celui où sont constatés le plus de décès. Il est de l’ordre de 70 ans dans la plupart des sociétés de chasseurs-cueilleurs.
Longevity among Hunter-Gatherers: A Cross-Cultural Examination [PDF]
Gurven & Kaplan
Population and development revue, 2007
–
High adult mortality among Hiwi hunter-gatherers: Implications for human evolution [PDF]
Hill et al.
Journal of human evolution, 2007
Here, we present mortality data for a fifth group, the Hiwi hunter-gatherers of Venezuela. The results show comparatively high death rates among the Hiwi and highlight differences in mortality rates among hunter-gatherer societies. The high levels of conspecific violence and adult mortality in the Hiwi may better represent Paleolithic human demographics than do the lower, disease-based death rates reported in the most frequently cited forager studies.
–
Older age becomes common late in human evolution [Texte]
Caspari & Lee
PNAS, 2004
Whereas there is significant increased longevity between all groups, indicating a trend of increased adult survivorship over the course of human evolution, there is a dramatic increase in longevity in the modern humans of the Early Upper Paleolithic. We believe that this great increase contributed to population expansions and cultural innovations associated with modernity.
–
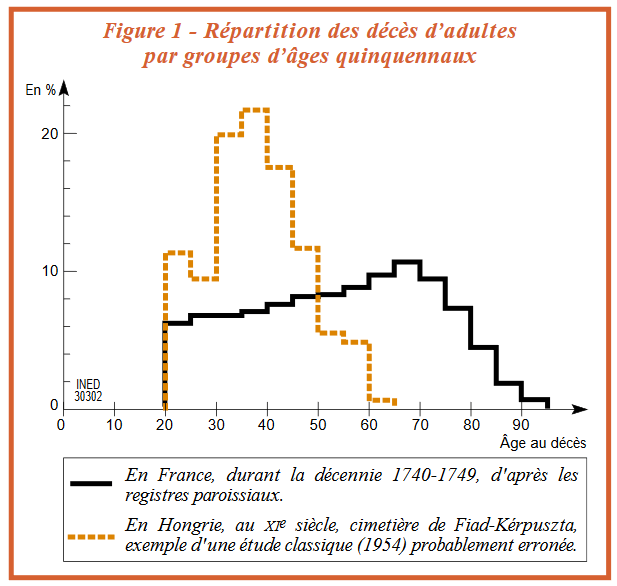
Certaines études un peu anciennes ont pu sous-estimer les âges au décès des adultes, pour des époques historiques, mais sans doute aussi préhistoriques :
A quel âge mouraient nos ancêtres ? [Texte]
Claude Masset
INED, Population et sociétés n°380, 2002
–
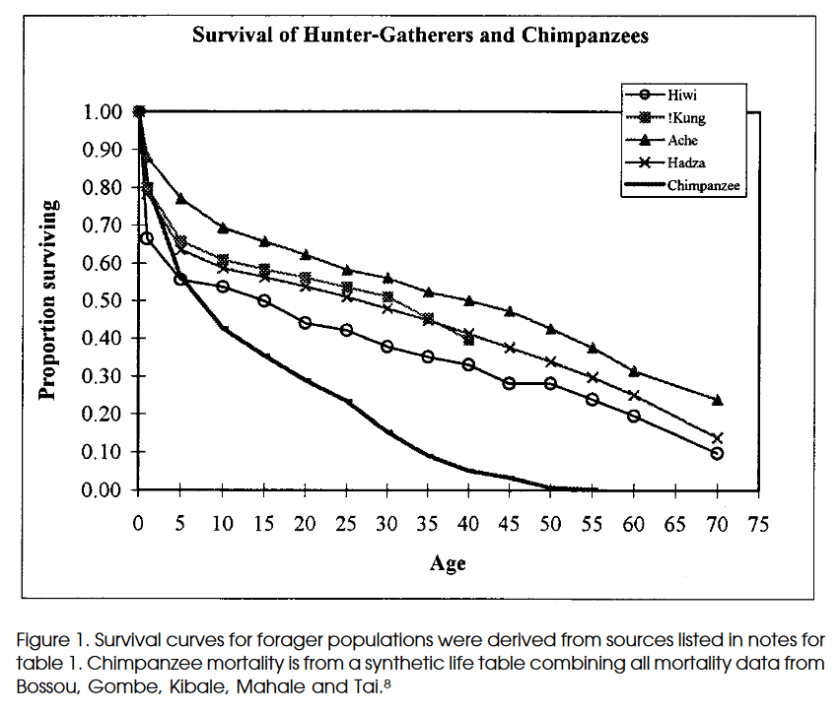
Ici, chez les Aché, environ 20% des individus atteignent l’âge de 70 ans, environ 10% pour les Hiwi.
A Theory of Human Life History Evolution: Diet, Intelligence, and Longevity [PDF]
Kaplan et al.
Evolutionnary anthropology, 2000
–
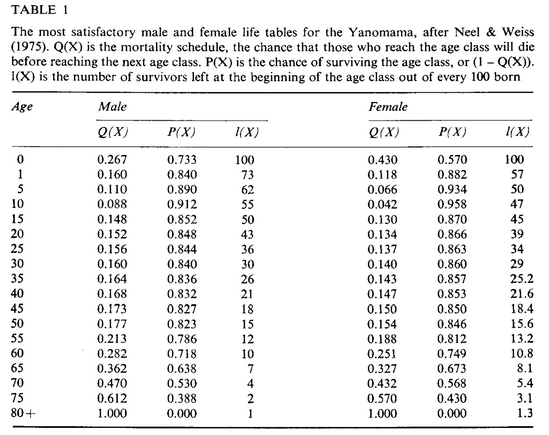
Cette étude chez les Yanomami rapporte environ 5% de survie à 70 ans et encore 1% à 80 ans.
The genetic structure of a tribal population, the Yanomama Indians [PDF]
Neel & Weiss
American journal of physical anthropology, 1975
–
Why Humans Live So Long [Texte]
Heather Pringle
Scientific American, 2015
Records de longévité contemporains
Evidence That Jeanne Calment Died in 1934—Not 1997
Nicolay Zak
Rejuvenation research, 2019
https://www.liebertpub.com/doi/full/10.1089/rej.2018.2167
I present a body of data that, I argue, cumulatively casts serious doubt on the validity of Jeanne Calment’s accepted world record of human life span. First, I assess the plausibility of the record based on the life spans of other centenarians in the International Database of Longevity (IDL) and critique some arguments put forward previously in support of that plausibility, including the longevity of Calment’s ancestors. Second, I review the literature dedicated to Calment and discuss multiple contradictions in her interviews, biographies, photos, and documents. I argue that the evidence from these sources motivates renewed consideration of the previously rejected hypothesis that Jeanne’s daughter Yvonne acquired her mother’s identity after her death to avoid financial problems and that Jeanne Calment’s death was reported as Yvonne’s death in 1934. Finally I discuss the importance of reconsidering the principles of validation, due to the possibility of similar problems regarding other exceptionally long-lived people and the mistaken inferences that researchers may draw from flawed datasets. The phenomenon of Jeanne Calment may prove to be an instructive example of the uncertainty of seemingly well-established facts.
The plateau of human mortality: demography of longevity pioneers
Barbi et al
Science, 2018
https://science.sciencemag.org/content/360/6396/1459
The increasing number of exceptionally long-lived people (Table 1) and the fact that their mortality beyond 105 is seen to be declining across cohorts—lowering the mortality plateau or postponing the age when it appears—strongly suggest that longevity is continuing to increase over time and that a limit, if any, has not been reached. Our results contribute to a recently rekindled debate (15–17) about the existence of a fixed maximum life span for humans, underwriting doubt that any limit is as yet in view.
Comment on “The plateau of human mortality: Demography of longevity pioneers”
Beltran-Sanchez et al.
Science, 2018
https://science.sciencemag.org/content/361/6409/eaav1200.full
Barbi et al. (Reports, 29 June 2018, p. 1459) reported that human mortality rate reached a “plateau” after the age of 105, suggesting there may be no limit to human longevity. We show, using their data, that potential lifespans cannot increase much beyond the current 122 years unless future biomedical advances alter the intrinsic rate of human aging.
Response to Comment on “The plateau of human mortality: Demography of longevity pioneers”
Barbi et al.
Science, 2018
https://science.sciencemag.org/content/362/6412/eaav3229.full
Beltrán-Sánchez et al. based their comment on misleading calculations of the maximum survival age. With realistic numbers of people attaining age 105 and the estimated plateau, the Jeanne Calment record is indeed plausible.
Errors as a primary cause of late-life mortality deceleration and plateaus
Saul Justin Newman
PLOS, 2018
https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.2006776
Here, it is shown that late-life mortality deceleration (LLMD) and late-life plateaus are caused by common demographic errors. […] ageing does not slow or stop during old age in humans, and that there is a finite limit to human longevity.
Hypothèse de la grand-mère
Déplacé sur [cette page]
Divers
Voyage en terres gérontocides : l’élimination des vieillards comme remède à la vieillesse ?
Nadine Bernard
Cahier des études anciennes, 2018
https://journals.openedition.org/etudesanciennes/1102
L’importance aux yeux des Grecs des notions d’âge et de classes d’âge conduit nos auteurs à poser un âge défini pour les exécutions, exprimé en multiples de 10 qui plus est, aussi surprenant soit-il pour des peuples jugés non civilisés et ignorants des computations calendaires15. De telles précisions traduisent des présupposés bien étranges pour les populations citées et renvoient plus sûrement aux déterminations de seuils fonctionnels ou physiologiques de la vieillesse en pays grec, à 60 ou 70 ans
The sex with the reduced sex chromosome dies earlier: a comparison across the tree of life [Texte]
Xirocostas et al.
Royal society biology letters, 2020
Surprisingly, we found substantial differences in lifespan dimorphism between female heterogametic species (in which the homogametic sex lives 7.1% longer) and male heterogametic species (in which the homogametic sex lives 20.9% longer). Our findings demonstrate the importance of considering chromosome morphology in addition to sexual selection and environment as potential drivers of sexual dimorphism
Il semble assez courant chez les transhumanistes de penser que la vie au paléolithique n’apportait aucun avantage évolutif après la naissance du dernier enfant. Au contraire, il leur semble avantageux pour les enfants que les parents décèdent rapidement, pour leur laisser des ressources. C’est très mal connaître l’organisation des sociétés primitives, le temps d’éducation nécessaire aux enfants, et l’importance de la survie des parents, voire des grands parents.
« L’inconséquence de ce qui nous arrive après l’âge de la reproduction n’avait guère d’importance quand on mourait peu d’années après la puberté ».
Laurent Alexandre, La mort de la mort, p44.