
L’évolution du cerveau humain pose une multitude de questions. Sachronologie, ses causes, ses facteurs limitants, ses conséquences…
Elle est liée notamment à la controverse « du feu ou de la viande » exposée dans cet article et les suivants, et plus succinctement dans ce thread twitter.
1. Données sur l’évolution du cerveau humain
2. Déterminants de l’évolution du cerveau
Données sur l’évolution du cerveau humain
L’évolution du cerveau est quantitative (un cerveau plus gros) et qualitative (la part relative de ses différentes parties évolue, des aires ou des réseaux différents s’organisent..).
Pour la partie quantitative, au moins deux aspects sont à considérer : la taille absolue du cerveau, et sa taille relative, avec la notion de coefficient d’encéphalisation.
Les méthodes employées sont diverses : mesure directe du volume intracrânien à partir des fossiles retrouvés, inférence de la forme du cerveau à partir des traces laisées à l’intérieur des boites crâniennes, ou mesure de la capacité d’irrigation sanguine de la boite crânienne…
An economic model and evidence of the evolution of human intelligence in the Middle Pleistocene: Climate change and assortative mating [Texte]
Bruce C. Petersen
PlosOne, 2023
When and Why Did Human Brains Decrease in Size? A New Change-Point Analysis and Insights From Brain Evolution in Ants [Texte]
Jeremy DaSilva
Frontiers in ecology and evolution, 2021
We find that hominin brains experienced positive rate changes at 2.1 and 1.5 million years ago, coincident with the early evolution of Homo and technological innovations evident in the archeological record. But we also find that human brain size reduction was surprisingly recent, occurring in the last 3,000 years.
–
The primitive brain of early Homo [PDF]
Ponce de Leon et al.
Science, 2021
Here, we show that the brains of early Homo from Africa and Western Asia (Dmanisi) retained a primitive, great ape–like organization of the frontal lobe. By contrast, African Homo younger than 1.5 million years ago, as well as all Southeast Asian Homo erectus, exhibited a more derived, humanlike brain organization. Frontal lobe reorganization, once considered a hallmark of earliest Homo in Africa, thus evolved comparatively late, and long after Homo first dispersed from Africa.
–
Divergence-time estimates for hominins provide insight into encephalization and body mass trends in human evolution [PDF]
Püschel et al.
Nature ecology & evolution, 2021
H. sapiens stands out due to its rapid increase in PEQ from its common ancestor with H. neanderthalensis (node 24), going from ~2.88 to 3.22 in ~0.55 Myr. That is 0.62 PEQ Myr–1, which is 1.68 times greater compared to the rate observed since the common ancestor of Homo (nodes 13/14 to 24) between 3.3 Ma and 0.6 Ma and 4.43 times greater in comparison to the rate observed since the common ancestor of G. gorilla and the ancestor of Homo (nodes 1 to 13/14) between 10.4 and 3.3 Ma.
Reconstructing the Neanderthal brain using computational anatomy [Texte]
Koshiyama et al.
Nature scientific reports, 2018
We found that early Homo sapiens had relatively larger cerebellar hemispheres but a smaller occipital region in the cerebrum than Neanderthals long before the time that Neanderthals disappeared. Further, using behavioural and structural imaging data of living humans, the abilities such as cognitive flexibility, attention, the language processing, episodic and working memory capacity were positively correlated with size-adjusted cerebellar volume. As the cerebellar hemispheres are structured as a large array of uniform neural modules, a larger cerebellum may possess a larger capacity for cognitive information processing. Such a neuroanatomical difference in the cerebellum may have caused important differences in cognitive and social abilities between the two species and might have contributed to the replacement of Neanderthals by early Homo sapiens.
Pattern and process in hominin brain size evolution are scale-dependent [PDF]
Du et al.
The Royal Society Publishing, 2018
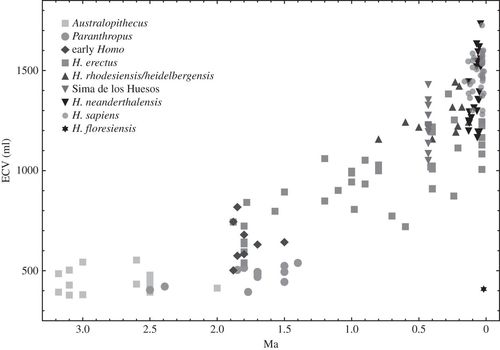
Our results show that fossil hominin ECV data at the clade level are most consistent with a gradual pattern of ECV increase through time. Understanding how this pattern emerged from processes that operate at lower taxonomic levels is more complicated. Our analyses are consistent with microevolutionary mechanisms as the dominant driver of clade-level change (64 or 88% of change using a more or less speciose taxonomy, respectively), alternating with secondary macroevolutionary mechanisms. This implies changing selective pressures and shifts in the relative importance of different evolutionary processes through time.
–
Les créationnistes aussi planchent sur l’évolution du cerveau :
Estimating the statistical significance of hominin encephalization [PDF]
TC Wood.
Journal of creation theology and science, 2016
Encephalization, the increase in endocranial volume through the Pleistocene, is examined from a creationist perspective. By correcting for body size using a linear regression model based on extant primates, the residual can be used to estimate the significance of the encephalization of fossil taxa. Using this method, significant encephalization is seen only in Homo and two australopiths, A. africanus and A. sediba. Since Wood (2010, 2016b) classifies all of Homo and A. sediba as human, these results indicate that all humans except H. naledi and only one nonhuman (A. africanus) are significantly encephalized. Based on these results, creationist neuroscientists ought to consider the possible causes of the encephalization trend in humans.
–
A paleoneurological survey of Homo erectus endocranial metrics [PDF]
Bruner et al.
Quaternary international, 2015
The taxonomic debate on the phylogenetic coherence of Homo erectus as a widespread intercontinental species is constantly put forward, without major agreements. Differences between the African and Asian fossil record as well as differences between the Chinese and Indonesian groups (or even within these two regions) have frequently been used to propose splitting taxonomical alternatives. In this paper, we analyze the endocranial variation of African and Asian specimens belonging to the hypodigm of H. erectus sensu lato, to assess whether or not these groups can be characterized in terms of traditional endocranialmetrics. According to the basic endocast proportions, the three geographic groups largely overlap in their phenotypic distribution and morphological patterns. The morphological affinity or differences among the specimens are largely based on brain size. As already evidenced by using other cranial features, traditional paleoneurological metrics cannot distinguish possible independent groups or trends within the Afro-Asiatic H. erectus hypodigm. Endocranial features and variability are discussed as to provide a general perspective on the paleoneurological traits of this taxon.
–
Brain ontogeny and life history in Pleistocene hominins [PDF]
Jean-Jacques Hublin et al.
The Royal Society Publishing, 2015
There is a consensus that Australopithecus endocasts show signs of brain reorganization and depart from the sulcal patterns found in apes, despite their ape-like endocranial volumes. It is therefore possible that brain reorganization in australopiths and its cognitive consequences underlie the subsequent brain expansion in the genus Homo. Furthermore, if supported by further investigations, the protracted pattern of brain development in A. afarensis would confirm that one cannot simply contrast a primitive ‘ape-pattern’ to a ‘human-pattern’.
–
[Hominisation par la chasse]
How humans evolved large brains: comparative evidence [PDF]
Isler & Van Schaik
Evolutionary anthropology, 2014
In conclusion, the archeological evidence for big-game hunting or scavenging at an early stage in the evolution of the genus Homo points towards a major role of cooperative hunting, which led to male food sharing, as a trigger of the remarkable increase in hominin brain size. Provisioning and babysitting by post-reproductive females and communal nursing among breeding females may have pre- or postdated this change in lifestyle, but from the comparative evidence we suspect that female help on its own would not have allowed for the evolution of the uniquely human combination of traits that arose between 2.5 and 2
million years ago.
–
Darwin’s mistake: Explaining the discontinuity between human and nonhuman minds [PDF]
Penn et al.
Behavioral and brain sciences, 2008
To wit, there is a significant discontinuity in the degree to which human and nonhuman animals are able to approximate the higher-order, systematic, relational capabilities of a physical symbol system (PSS) (Newell 1980). We show that this symbolic-relational discontinuity pervades nearly every domain of cognition and runs much deeper than even the spectacular scaffolding provided by language or culture alone can explain. We propose a representational-level specification as to where human and nonhuman animals’ abilities to approximate a PSS are similar and where they differ. We conclude by suggesting that recent symbolic-connectionist models of cognition shed new light on the mechanisms that underlie the gap between human and nonhuman minds.
–
Humans Have Evolved Specialized Skills of Social Cognition: The Cultural Intelligence Hypothesis [PDF]
Herrmann et al.
Science, 2007
Supporting the cultural intelligence hypothesis and contradicting the hypothesis that humans simply have more “general intelligence,” we found that the children and chimpanzees had very similar cognitive skills for dealing with the physical world but that the children had more sophisticated cognitive skills than either of the ape species for dealing with the social world.
–
Body size, body proportions, and encephalization in a Middle Pleistocene archaic human from northern China [PDF]
Rosenberg et al.
PNAS, 2006

–
Scaling of brain and body weight within modern and fossil hominids: implications for the Flores specimen [PDF]
Schoenemann & Allen
American journal of physical anthropology, 2006
The relationship between encephalization quotient (EQ) and body weight is found to be consistently negative within all hominid species
–
Early brain growth in Homo erectus and implications for cognitive ability [PDF]
Coqueugniot et al. (Hublin)
Nature, 2004
Here we present the results of an analysis of the 1.8-million-year-old Mojokerto child (Perning 1, Java), the only well preserved skull of a Homo erectus infant, by computed tomography. Comparison with a large series of extant humans and chimpanzees indicates that this individual was about 1 yr (0–1.5 yr) old at death and had an endocranial capacity at 72–84% of an average adult H. erectus. This pattern of relative brain growth resembles that of living apes, but differs from that seen in extant humans. It implies that major differences in the development of cognitive capabilities existed between H. erectus and anatomically modern humans.
–
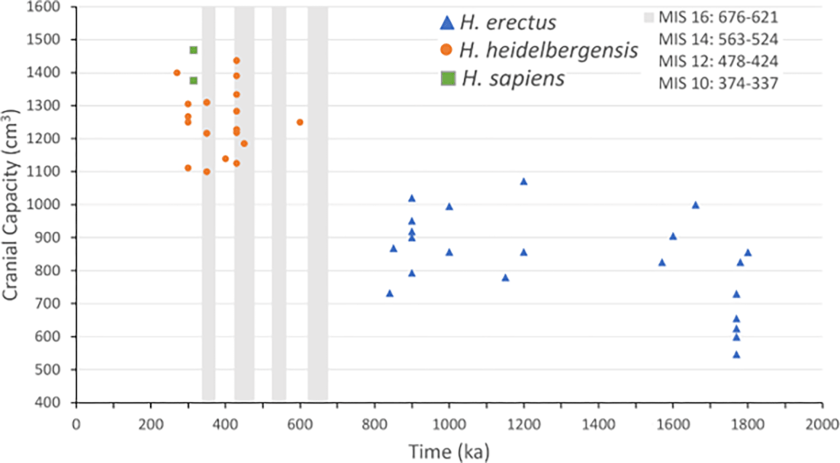
Suggère une première augmentation significative de l’encéphalisation avant -1,8Ma suivie d’un plateau, puis d’une seconde augmentation significative entre -600 et -150ka. L’une initiée par le passage à l’alimentation carnée, l’autre permise par le feu ?
Body mass and encephalization in pleistocene Homo [Abstract]
Ruff et al.
Nature, 1997
On the basis of an analysis of 163 individuals, body mass in Pleistocene Homo averaged significantly (about 10%) larger than a representative sample of living humans. Relative to body mass, brain mass in late archaic H. sapiens (Neanderthals) was slightly smaller than in early ‘anatomically modern’ humans, but the major increase in encephalization within Homo occurred earlier during the Middle Pleistocene (600–150 thousand years before present (kyr BP)), preceded by a long period of stasis extending through the Early Pleistocene (1,800 kyr BP).
Déterminants de l’évolution du cerveau
Prey Size Decline as a Unifying Ecological Selecting Agent in Pleistocene Human Evolution [Texte]
Ben-Dor & Barkai
Quaternary, 2021
We hypothesize that megafauna extinctions throughout the Pleistocene, that led to a progressive decline in large prey availability, were a primary selecting agent in key evolutionary and cultural changes in human prehistory. The Pleistocene human past is characterized by a series of transformations that include the evolution of new physiological traits and the adoption, assimilation, and replacement of cultural and behavioral patterns. Some changes, such as brain expansion, use of fire, developments in stone-tool technologies, or the scale of resource intensification, were uncharacteristically progressive.
–
Evolution of the Human Brain: the key roles of DHA (omega-3 fatty acid) and Δ6-desaturase gene [PDF]
How scientists perceive the evolutionary origin of human traits: Results of a survey study [Texte]
Tuomisto et al.
Ecology and evolution, 2018
The hypotheses proposing that encephalization was triggered by improved nutrition also received intermediate popularity scores, whether achieved by cooking or by increased consumption of fish or meat (all three with credibility scores in the range 2.61–2.77).
The evolutionary roles of nutrition selection and dietary quality in the human brain size and encephalization [Texte]
Burini & Leonard
Nutrire, 2018
In addition to the energetic benefits associated with greater meat consumption, it appears that such a dietary shift would have also provided increased levels of key fatty acids necessary for supporting the rapid hominid brain evolution [60].
Half of human brain composition is fat, and 20% of its dry weight is long-chain polyunsaturated fatty acids (LCPUFAs). Consequently, improvements in consumption of dietary fat were a necessary condition for promoting encephalization [61, 62].
Mammalian brain growth is dependent upon sufficient amounts of two LCPUFAs: docosahexaenoic acid (DHA) and arachidonic acid (AA), and it appears that mammals have a limited capacity to synthesize these fatty acids from dietary precursors. Hence, species with higher levels of encephalization would have greater requirements for DHA and AA [62]. Consequently, dietary sources of DHA and AA were likely limiting nutrients that constrained the evolution of larger brain size in many mammalian lineages [63].
Meat and Nicotinamide: A Causal Role in Human Evolution, History, and Demographics [PDF]
Williams & Hill
International Journal of Tryptophan Research, 2017
–
Control of Fire in the Paleolithic: Evaluating the Cooking Hypothesis [Texte]
Richard Wrangham
Current Anthropology, 2017
–
DeCasien et al.
Nature ecology & evolution, 2017
Here, we use a much larger sample of primates, more recent phylogenies, and updated statistical techniques, to show that brain size is predicted by diet, rather than multiple measures of sociality, after controlling for body size and phylogeny. Specifically, frugivores exhibit larger brains than folivores. Our results call into question the current emphasis on social rather than ecological explanations for the evolution of large brains in primates and evoke a range of ecological and developmental hypotheses centred on frugivory, including spatial information storage, extractive foraging and overcoming metabolic constraints.
Mais attention, ce résultat ne peut pas être étendu aux humains, dont le cerveau est trop atypique :

–
Human Brain Expansion during Evolution Is Independent of Fire Control and Cooking [Texte]
Alianda M. Cornelio et al.
Frontiers in neurosciences, 2016
–
Brain size and thermoregulation during the evolution of the genus Homo [PDF]
Naya et al.
Comparative biochemistry and physiology, 2016
–
Metabolic costs and evolutionnary implications of human brain development [Texte]
Kuzawa et al.
Proceedings of the national academy of science, 2014
We find that the brain’s metabolic requirements peak in childhood, when it uses glucose at a rate equivalent to 66% of the body’s resting metabolism and 43% of the body’s daily energy requirement, and that brain glucose demand relates inversely to body growth from infancy to puberty. Our findings support the hypothesis that the unusually high costs of human brain development require a compensatory slowingof childhood body growth.
–
Earliest fire in Africa: towards the convergence of archaeological evidence and the cooking hypothesis [PDF]
Gowlett & Wrangham
Azania: Archaeological research in Africa, 2013
–
Big Brains, Meat, Tuberculosis, and the Nicotinamide Switches: Co-Evolutionary Relationships with Modern Repercussions? [Texte] [PDF]
Williams & Dunbar
International journal of tryptophan research, 2013
Meat-eating was a game changer for human evolution. We suggest that the limiting factors for expanding brains earlier were scarcities of nicotinamide and tryptophan. In humans and some other omnivores, lack of meat causes these deficiencies. Nicotinamide adenine dinucleotide (NADH) is necessary to synthesize adenosine triphosphate (ATP) via either glycolysis or via the mitochondrial respiratory chain. NAD consumption is also necessary for developmental and repair circuits. Inadequate supplies result in “de-evolutionary” brain atrophy, as seen with pellagra.
–
Docosahexaenoic Acid and Shore-Based Diets in Hominin Encephalization: A Rebuttal [PDF]
Cunnane et al.
American journal of human biology, 2007
Meat made us Human, In Evolution of the Human Diet: The Known, the Unknown, and the Unknowable (Livre) [Google Books]
Henry T. Bunn
Oxford University Press, 2006
–
The Critical Role Played by Animal Source Foods in Human (Homo) Evolution [Texte]
Katharine Milton
The Journal of Nutritin, 2003
–
Survival of the fattest: fat babies were the key to evolution of the large human brain [PDF]
Cunnane & Crawford
Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2003
–
Metabolic correlates of hominid brain evolution [Abstract] [PDF]
Leonard et al.
Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2003
–
Adipose tissue in human infancy and childhood: An evolutionary perspective [PDF]
Christopher Kuzawa
American Journal of biological anthropology, 1999
the greater adiposity of human neonates is at least partially explainable as an accompaniment of the enlarged human brain, which demands a larger energy reserve to ensure that its obligatory needs are met when the flow of resources from mother or other caretakers is disrupted.
–
Animal Fat and Cholesterol May Have Helped Primitive Man Evolve a Large Brain [Abstract]
Franck D. Mann
Perspectives in biology and medicine, 1998
–
The possible role of long-chain, omega-3 fatty acids in human brain phylogeny [Abstract]
Chamberlain
Perspectives in biology and medicine, 1996
The expensive-tissue hypothesis : the brain and the digestive system in human and primate evolution [PDF]
Aiello & Wheeler
Current Anthropology, 1995.
–
Sur le rôle de l’alimentation dans l’évolution du cerveau, mais ici, plutôt les fruits, et pour les primates non humains (les humains sont exclus parce que la taille de leur cerveau est trop atypique) :
Primate brain size is predicted by diet but not sociality [PDF]
DeCasien et al.
Nature ecology & evolution
Articles et blogs
Article de Miki Ben-Dor tiré de sa thèse :
Discussing The Cooking Hypothesis – a section from my Ph.D thesis [Texte]
Défense de la Cooking Hypothesis :
Earlier Evidence for Erectus’ Use of Fire [Texte]
Visuels
Graphique évolution du cerveau et acquisitions culturelles humaines
Bradshaw foundation, 2012, pour le visuel original.
http://bradshawfoundation.com/origins/index.php

