
La discussion sur l’évolution du dimorphisme humain depuis le pliocène n’est sans doute pas terminée. Cependant, les publications des 10 dernières années semblent plaider pour un dimorphisme important il y a 3 millions d’années, ayant fortement décru jusqu’à Homo erectus, plus faiblement ensuite. Il est possible qu’il y ait une stabilisation, et pourquoi pas une légère réaugmentation du dimorphisme au sein d’Homo sapiens, mais les données sont parfois contradictoires, et le dimorphisme chez Homo sapiens resterait en tous les cas faible par rapport à ses prédécesseurs, sous réserve que les méthodes récentes de calcul et de diagnose des sexes soient désormais suffisamment fiables.
Pour les autres manifestations du dimorphisme que la taille et le poids (musculature, zone pelvienne, latéralisation, voix, etc…) [Page]
1. Évolution du dimorphisme jusqu’à Sapiens
2. Dimorphisme chez néandertal
3. Évolution du dimorphisme au sein d’Homo sapiens
4. Divers
Evolution du dimorphisme sexuel jusqu’à Sapiens
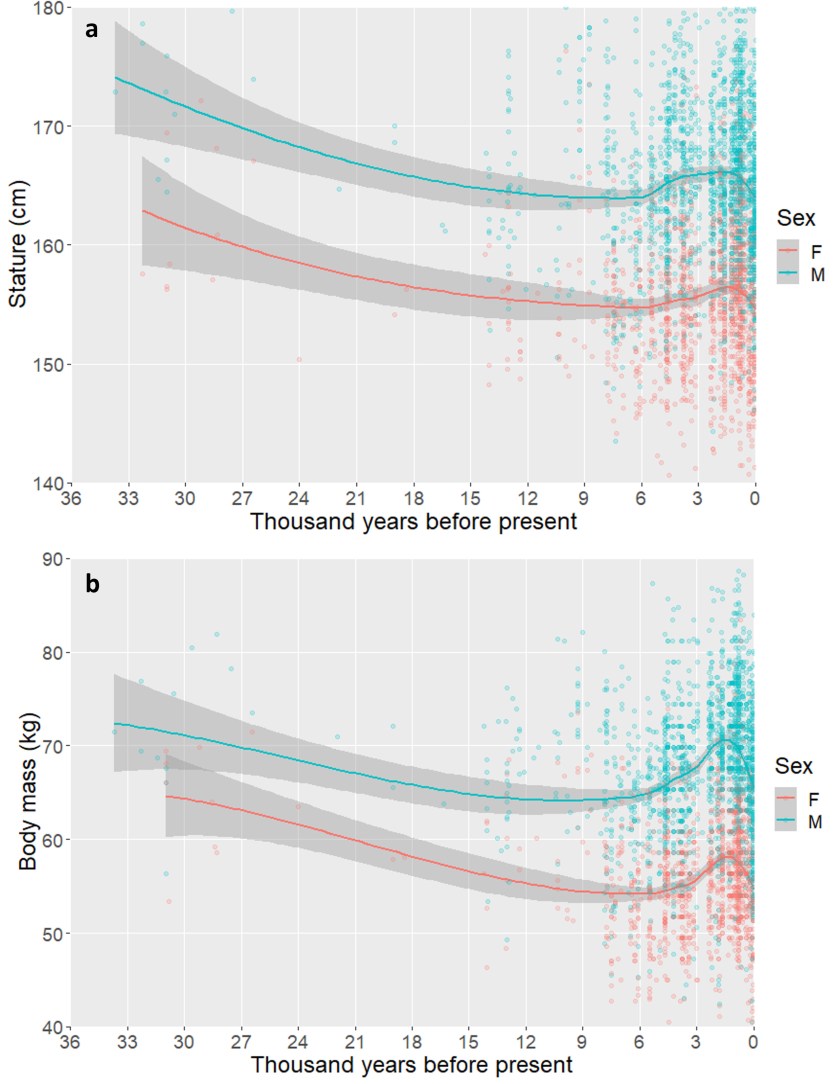
Long-term trends in human body size track regional variation in subsistence transitions and growth acceleration linked to dairying [Texte]
Stock et al. (Ruff)
PNAS, 2023
Our results demonstrate that 1) decreases in stature preceded the origins of agriculture in some regions; 2) the Levant and China, regions of in situ domestication of species and an extended period of mixed foraging and agricultural subsistence, had stable stature and body mass over time; and 3) stature and body mass increases in Central and Northern Europe coincide with the timing of selective sweeps for lactase persistence, providing support for the “Lactase Growth Hypothesis.”
–
The Australopithecus assemblage from Sterkfontein Member 4 (South Africa) and the concept of variation in palaeontology [Texte]
Amélie Beaudet
Evolutionary Anthropology, 2023
–
Sexual dimorphism in Homo erectus inferred from 1.5 Ma footprints near Ileret, Kenya [Texte]
Villmoare et al.
Nature scientific reports, 2019
Our results indicate that East African Homo erectus was more dimorphic than modern Homo sapiens, although less so than highly dimorphic apes, suggesting that the Ileret footprints offer a unique window into an important transitional period in hominin social behavior.
–
Sexual Dimorphism in Hominin Ancestors [Abstract]
J. Michael Plavcan
The international encyclopedia of anthropology, 2018
[…] strong dimorphism in Australopithecus suggests polygynous mating, with strong male competition. Changes in dimorphism and body size suggest changes in female life history and associated patterns of male competition. Finally, the reevolution of dimorphism in hominins suggests that the modern human condition is derived and not inherited from the common ancestor of Homo and Pan.
–
Lower limb articular scaling and body mass estimation in Pliocene and Pleistocene hominins [PDF]
Ruff et al.
Journal of human evolution, 2018
Sexual dimorphism in body mass is greatest in Australopithecus afarensis (log[male/female] = 1.54), declines in Australopithecus africanus and Paranthropus robustus (log ratio 1.36), and then again in early Homo and middle and late Pleistocene archaic Homo (log ratio 1.20–1.27), although it remains somewhat elevated above that of living and middle/late Pleistocene anatomically modern humans (log ratio about 1.15).
–
Long-term patterns of body mass and stature evolution within the hominin lineage [Texte]
Will et al.
Royal society open science, 2017
our taxonomic analyses show that among Homo, younger species tend to be less variable in body size than older taxa (figure 8d), potentially associated with decreasing sexual dimorphism
–
New footprints from Laetoli (Tanzania) provide evidence for marked body size variation in early hominins [Texte]
Masao et al.
Elife, 2016
Our results are consistent with considerable body size variation and, probably, degree of sexual dimorphism within a single species of bipedal hominins as early as 3.66 million years ago.
–
Evolutionary perspectives on human height variation [Abstract]
Stulp & Barrett
Biological reviews, 2016
Here, we present a synthetic review of the literature on human height from an explicit evolutionary perspective, addressing its phylogenetic history, development, and environmental and genetic influences on growth and stature. In addition to presenting evidence to suggest the past action of natural selection on human height, we also assess the evidence that natural and sexual selection continues to act on height in contemporary populations. Although there is clear evidence to suggest that selection acts on height, mainly through life‐history processes but perhaps also directly, it is also apparent that methodological factors reduce the confidence with which such inferences can be drawn, and there remain surprising gaps in our knowledge. The inability to draw firm conclusions about the adaptiveness of such a highly visible and easily measured trait suggests we should show an appropriate degree of caution when dealing with other human traits in evolutionary perspective.
–
Body mass estimates of hominin fossils and the evolution of human
body size [PDF]
Grabowski et al.
Journal of human evolution, 2015

Soit Afarensis : 1,59 ; Early Homo : 1,42 ; Erectus : 1,17 ; Sapiens : 1,12 ; Pan troglodytes (chimpanzé) : 1,30
–
Evolution of early Homo: An integrated biological perspective [PDF]
Anton et al.
Science, 2014

–
Body Size, Size Variation, and Sexual Size Dimorphism in Early Homo [PDF]
J. Michael Plavcan
Current anthropology, 2012
Modern humans show an unusual pattern of relatively modest body mass dimorphism (about 15%; Smith and Jungers 1997) but moderate “lean” body mass dimorphism (about 44%; Wang et al. 2001). This latter value reflects a proportionally greater amount of fat in human females (Plavcan 2012; Wang et al. 2001; Wells 2007, 2012). Skeletal dimorphism in modern humans is moderate and slightly greater than that of chimpanzees (Gordon, Green, and Richmond 2008), but when corrected for dimensionality it is proportional to lean body mass dimorphism in magnitude (Plavcan 2012). Cranial dimorphism in modern humans tends to be modest, being intermediate between body mass dimorphism and lean body mass dimorphism (Plavcan 2012).
Because most cranial, dental, and postcranial traits are normally or at least unimodally distributed in males and females, the effect of dimorphism is to simultaneously increase the total sample variance and generate bimodality
–
Body size, body proportions, and encephalization in a Middle Pleistocene archaic human from northern China [PDF]
Rosenberg et al.
PNAS, 2006

–
Sexual dimorphism in Australopithecus afarensis revisited: How strong is the case for a human-like pattern of dimorphism ? [PDF]
Plavcan et al.
Journal of human evolution, 2005
we herein highlight several shortcomings in the analysis of Reno et al. (2003) that strongly impact the conclusion that A. afarensis showed a human-like level of sexual size dimorphism.
–
Sexual dimorphism in Australopithecus afarensis was similar to that of modern humans [PDF]
Reno et al
PNAS, 2003
skeletal size dimorphism in A. afarensis was most similar to that of contemporary Homo sapiens. These data eliminate some apparent discrepancies between the canine and skeletal size dimorphism in hominoids, imply that the species was not characterized by substantial sexual bimaturation, and greatly increase the probability that the reproductive strategy of A. afarensis was principally monogamy.
–
Variation in human body size and shape [PDF]
Christopher Ruff
Annual review of anthropology, 2002

–
Human evolution : taxonomy and paleobiology [PDF]
Wood & Richmond
The Journal of anatomy, 2000
Variation in body size suggests that, by the
Middle Pleistocene, sexual size dimorphism was
comparable to that of modern humans (Arsuaga et al.
1997 a ; Quinney & Collard, 1997).
–
Randomization procedures and sexual dimorphism in Australopithecus afarensis
Lockwood et al.
Journal of Human Evolution, 1996
The estimate of average sexual dimorphism for mandibular size in A. afarensis, based on the CV of geometric means, gives a result (ISD=1·167) that is compatible with a single species that possesses a relatively high degree of sexual dimorphism
Dimorphisme sexuel chez néandertal
Sexual Dimorphism in the Mandible of Homo Neanderthalensis and Homo Sapiens: Morphological Patterns and Behavioural Implications [PDF]
Quiney & Collard
Oxbow books, 1997
In conclusion, we find that using mandibular measurements as a proxy of overall sexual dimorphism indicates that the Neanderthals exhibited a level of dimorphism that is indistinguishable from that of recent humans.
–
Size Variation in Middle Pleistocene Humans [PDF]
Arsuaga et al.
Science, 1997
It has been suggested that European Middle Pleistocene humans, Neandertals, and prehistoric modern humans had a greater sexual dimorphism than modern humans. Analysis of body size variation and cranial capacity variation in the large sample from the Sima de los Huesos site in Spain showed instead that the sexual dimorphism is comparable in Middle Pleistocene and modern populations.
–
Sexual differences in Neanderthal limb bones[Abstract]
Eric Trinkaus
Journal of human evolution, 1980
Neanderthal sexual size dimorphism, both within single site samples and in the total sexable sample, is virtually the same as that of recent human samples. Furthermore, despite a tendency towards more robust limbs, the Neanderthals exhibit sexual dimorphism in limb bone shaft and articular robusticity similar to that of recent human samples. By the time of the Neanderthals, sexual dimorphism in limb bone size and robusticity appears to have reached recent human proportions.
Evolution du dimorphisme sexuel au sein d’Homo sapiens
Sexual dimorphism [PDF]
Berner et al.
In Skeletal Variation and Adaptation in Europeans: Upper Paleolithic to the Twentieth Century, Christopher Ruff, 2018
As expected, male body size was larger overall and in almost all temporal subgroups. Average SD in body mass (17%) and stature (6%) are similar to means reported for modern humans generally (Stini, 1976; Gray and Wolfe, 1980; Ruff, 2002; Garvin, 2012). Our results showed essentially no change in SD in stature, and only a slight decrease in SD of body mass between the Upper Paleolithic and Mesolithic periods, followed by a more marked increase in the Neolithic period.
[…] In our sample, both males and females declined in body size following the UP
[…] We found a small increase in SD of stature and a larger increase in SD of body mass during the transition from the Mesolithic to Neolithic, resulting from an increase in male average stature and a decrease in female average body mass.
[…] evidence for consistent effects of agricultural versus foraging subsistence strategies on stature SD in living human populations is mixed. Except for body mass in females, our results do not indicate a decline in body size with the introduction of agriculture in Europe, nor a decline in SD of body size. Neither observation supports a general scenario of decreased health (at least as reflected in growth in body size) at the introduction of food production in Europe
[…] Nor is there any evidence in our data for significant changes in body size SD during agricultural intensification in the Bronze Age, although there is a slight decline in the Iron Age/Roman period.
[…] Males decline in body size more than females in the early modern period, then increase more in the recent period, leading
to a reduction followed by an increase in SD.
–
Temporal and Geographic Variation in Body Size and Shape of Europeans from the Late Pleistocene to Recent Times [Abstract]
Niskanen et al.
In Skeletal Variation and Adaptation in Europeans: Upper Paleolithic to the Twentieth Century, Christopher Ruff, 2018

–
The paleobiology of two adult skeletons from Baousso da Torre (Bausu da Ture) (Liguria, Italy): Implications for Gravettian lifestyle [Texte]
Villotte et al.
Comptes rendus Palevol, 2017
–
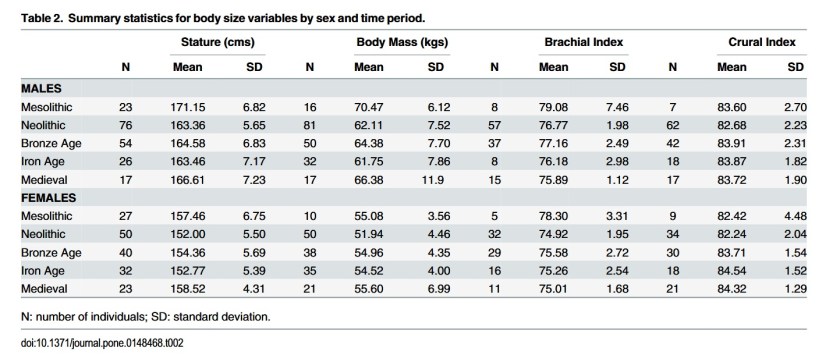
Early Life Conditions and Physiological Stress following the Transition to Farming in Central/Southeast Europe: Skeletal Growth Impairment and 6000 Years of Gradual Recovery [PDF]
Macintosh et al.
PLOS One, 2016
Results document significantly reduced mean stature, body mass, and crural index in Neolithic agriculturalists relative both to Late Mesolithic hunter-gatherer-fishers and to later farming populations.This indication of relative growth impairment in the Neolithic, particularly among women, is supported by existing evidence of high developmental stress, intensive physical activity, and variable access to animal protein in these early agricultural populations. Among subsequent agriculturalists, temporal increases in mean stature, body mass, and crural index were more pronounced among Central European women, driving declines in the magnitude of sexual dimorphism through time. Overall, results suggest that the transition to agriculture in Central/Southeast Europe was challenging for early farming populations, but was fol-lowed by gradual amelioration across thousands of years, particularly among Central European women. This sex difference may be indicative, in part, of greater temporal variation in the social status afforded to young girls, in their access to resources during growth, and/or in their health status than was experienced by men.
–
Morphological variation in Homo erectus and the origins of developmental plasticity [Page]
Anton et al.
Philosophical transactions of the Royal Society B, 2016

–
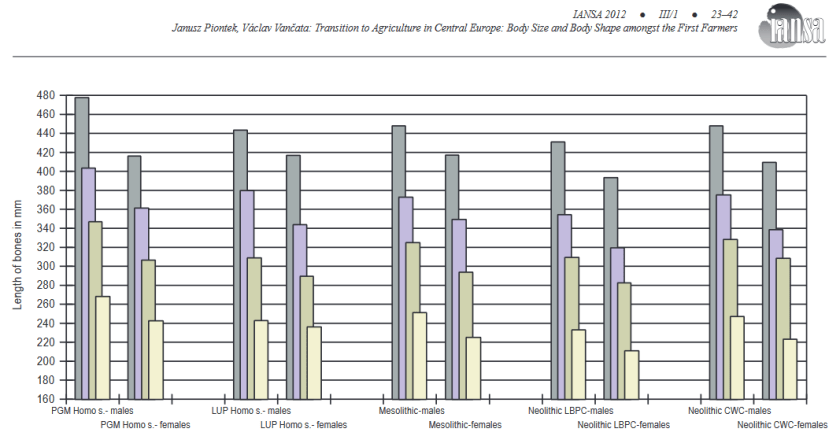
Transition to agriculture in central europe: Body size and Body shape amongst the First Farmers [PDF]
Piontek & Vancata
Interdisciplinaria archeologica, 2012
PGM = pre-glacial maximum (aurignacien/gravettien
–
Hunters of the Ice Age: The biology of Upper Paleolithic people [PDF]
Holt & Formicola
American journal of anthropology, 2008

–
Stature of early Europeans [PDF]
Michael Hermanussen
Hormones, 2003
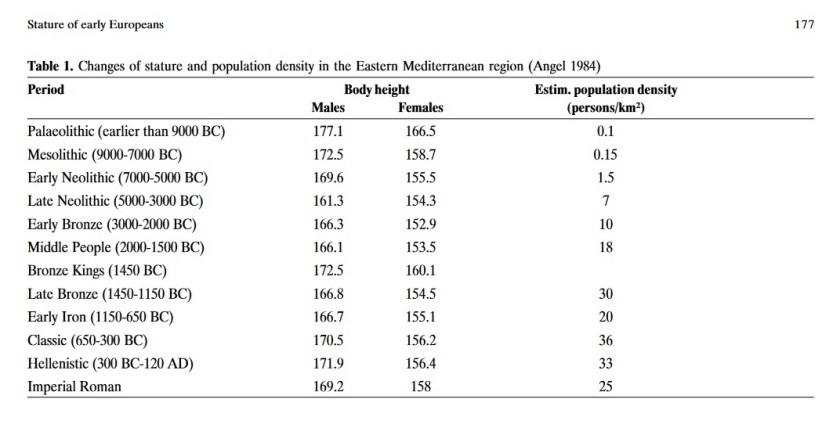
–
Sexual dimorphism in human lower limb bone structure: relationship to subsistence strategy and sexual division of labor [Abstract]
Christopher Ruff
Journal of Human evolution, 1987
There is a consistent decline in sexual dimorphism from hunting-gathering to agricultural to industrial subsistence strategy levels in properties which measure relative anteroposterior bending strength of the femur and tibia in the region about the knee. This trend parallels and is indicative of reductions in the sexual division of labor, in particular differences in the relative mobility of males and females. Sexual dimorphism in mediolateral bending strength near the hip shows no consistent temporal trend, probably reflecting relatively constant sex differences in pelvic structure related to the requirements of childbirth. Upper and Middle Paleolithic samples are indistinguishable in terms of sexual dimorphism from modern hunter-gatherers, suggesting a similar sexual division of labor.
–
(Cité par Priscille Touraille)
Sexual dimorphism and cultural evolution in the Late Pleistocene and Holocene of Europe [Abstract] [PDF]
David W. Frayer
Journal of Human Evolution, 1980
Dental, cranial and body size data are reviewed for European Upper Paleolithic, Mesolithic and Neolithic males and females. Over these three periods there is a substantial decrease in the level of sexual dimorphism. From separate analysis of trends occurring between males and females, it is shown that the major cause for this decrease in sexual dimorphism is gracilization of the males between the Upper Paleolithic and Mesolithic. Reduction in males is related to shifting technological patterns associated with hunting and changes in the types of animals hunted. Further reduction in sexual dimorphism between the Mesolithic and Neolithic and from the Neolithic to modern European populations is shown to be more closely tied to changes occurring among females. Analysis of changing patterns of sexual dimorphism in Late Pleistocene and Holocene populations of Europe suggests an interrelationship between cultural and biological evolution.
–
Sexual dimorphism and human tooth size differences [Abstract] [PDF]
Brace & Ryan
Journal of Human Evolution, 1980
The conclusion offered is that the smallest degree of sexual dimorphism visible in the modern world is to be found among those populations that are separated by the greatest interval of time from precursors who depended for their survival on a Pleistocene big game hunting mode of subsistence.
Divers
Sexual Size Dimorphism, Canine Dimorphism, and Male-Male Competition in Primates [Abstract]
J. Michael Plavcan
Human nature, 2012
What Are the Causes of Size Dimorphism in Modern Humans? The evolutionary origins of the prominent secondary sexual characteristics of humans are potentially diverse. Sexual ornamentation and size dimorphism in humans are hypothesized to have evolved for signaling to attract mates, to indicate mate quality(genetic quality in both sexes, and fecundity in particular in females) or age, to signal dominance, or even to reduce a sense of threat (Sefcek et al.2006). The greater body size and strength of males has been thought to give males competitive advantage for access to mates, or to be attractive to females, to signal male quality (Cashdan1996;Dixson2009; Sefcek et al.2006), or to reflect the sexual division of labor for hunting or between-group competition (Frayer1980). Body mass dimorphism also might reflect smaller female body size selected to increase or decrease female fecundity(Dixson2009; Ralls1976). Alternatively, human size and stature dimorphism may be a product of sexual selection from the recent past that persists in modern humans(Dixson2009; Rogers and Mukherjee1992; Wade and Shuster2004).
–
Short stature in African pygmies is not explained by sexual selection [Abstract]
Becker et al. (Priscille Touraille)
Evolution and human behaviour, 2012
Altogether, our results demonstrate that stature influences mate choice in pygmies, and we argue that, if of any influence for sexual selection, mate choice should have favored tallness rather than shortness in our pygmy population. Consequently, this study establishes that sexual selection is a very unlikely candidate to account for the evolution of pygmies’ short stature.
–
Why are women smaller than men? When anthropology meets evolutionary biology [PDF]
Touraille & Gouyon
Nature preceedings, 2008 (non reviewé)
There are large variations of size among humans but in all populations, men are larger on average than women. For most biologists this fact can be easily explained by the same processes that explain the size dimorphism in large mammals in general and in apes in particular. Due to fights between males for the possession of females, sexual selection has favoured bigger males. Indeed, this factor certainly explains why males are selected for being large but lets aside the question of selection on the female side. Actually, it has been shown that larger females are also favoured by natural selection. This is particularly relevant for women because their probability of dying when giving birth is then reduced. In this paper, the common view that size dimorphism in humans results from the fact that the advantage of being big is stronger for men than for women is challenged by another hypothesis, namely that the difference results from a difference of cost rather than from a difference of benefits. The cost of being big would be higher in women simply because, under gender hierarchical regimes found in all cultures, men are allocated the best food. The interaction between evolutionary forces and cultural practices could then lead to this disadaptive situation
–
Une espèce très dimorphique ou deux espèces différentes ? Une des grosses difficultés quand on travaille sur des fossiles rares, anciens et incomplets.
Size variation and sexual dimorphism in Australopithecus afarensis and living hominoids [Abstract]
Richmond & Jungers
Journal of human evolution, 1995
If the fossils from Hadar and Maka (and Laetoli) are assumed instead to be from one sexually dimorphic species, then the degree of sexual dimorphism of A. afarensis would have been at least as extreme as that of the most dimorphic living apes, the gorilla and orang-utan. It follows that a strictly monogamous social structure would have been highly unlikely.
–
(cité par Priscille Touraille)
Size, Sexual Dimorphism, and Polygyny in Primates, in Size and Scaling in Primate Biology (chapitre de livre)
Among primates, the extent of sexual dimorphism in body size ranges from species where mature females are slightly larger than mature males, as in some of the marmosets and tamarins (Ralls, 1976), through species where males are slightly larger than females, like many of the diurnal lemurs and the arboreal colobines, to those where males are nearly twice as heavy as females, as in the larger cercopithecines, the gorilla and the orang (Clutton-Brock and Harvey, 1978).
–
Subsistence practices and human sexual dimorphism of stature [Abstract] [PDF]
Wolfe & Gray
Journal of Human evolution, 1982
Hypotheses recently advanced by Brace & Ryan (1980) and Frayer (1980) suggest links between changes in human sexual dimorphism and changes in technology and subsistence practices. In this paper we test these hypotheses using a sample of extant human groups. Results indicate that extant agriculturalists exhibit a greater degree of sexual dimorphism in stature than extant hunter-gatherers. Moreover, the data analysed in this paper do not indicate that a more equal division of labor is associated with a decrease in human height sexual dimorphism.
