

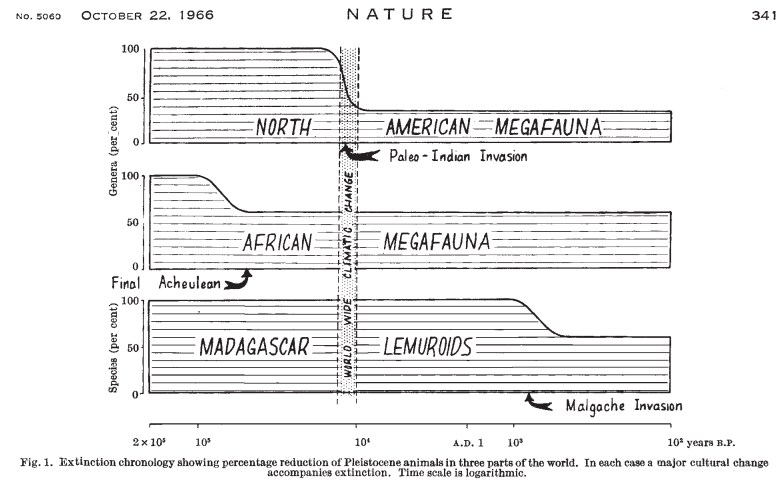
Le paléolithique supérieur a vu dans un laps de temps relativement court la disparition de nombreuses espèces, et notamment, en Eurasie, Amériques et Australie, une diminution massive de la mégafaune, les plus gros animaux. Certaines études suggèrent que cette diminition massive aurait commencé bien plus tôt, en Afrique, il y a peut-être plus de 2 millions d’années.
Les causes de ces disparitions sont âprement disputées, pour des raisons techniques, d’abord (il est compliqué de démêler les indices de ces périodes anciennes), mais aussi pour les conclusions qu’il est possible d’en tirer sur la « nature » humaine (voir dernière section).
1. Etudes générales
2. Etudes locales
3. Extinction de Neandertal [Déplacé ici]
4. Controverses dans les médias et les blogs
Etudes générales
Stewart et al., The state of the late Quaternary megafauna extinction debate: a systematic review and analysis, Frontiers in Mammal Science, 2025
« Overall, the major extinction hypotheses are similarly represented in our literature sample: 82 (23%) articles cite humans as the primary driver, 82 (23%) articles cite climate as the primary driver, 71 (20%) proposed a mixed human-climate cause. A third of articles offered no explicit major driver of extinction, while only a few considered an extraterrestrial cause, such as a solar flare or comet impact.«
Hauffe et al., Trait-mediated speciation and human-driven extinctions in proboscideans revealed by unsupervised Bayesian neural networks, ScienceAdvances, 2024 [Texte]
« We estimated the proboscideans extinction rates to be most strongly affected by the overlap with humans, followed by more limited effects of geographic distribution and ecomorphology linked with tusk and mandible shapes (Fig. 4C and table S2). On the basis of partial dependent plots, the spatiotemporal overlap with early humans starting around 1.8 Ma was associated with a 5-fold increase in extinction rate, while the effect of modern Homo sapiens in the Late Pleistocene and Holocene was linked with a 17-fold increase.«
Svenning et al., The late-Quaternary megafauna extinctions: patterns, causes, ecological consequences, and implications for ecosystem management in the Anthropocene, Cambridge Prisms: Extinctions, 2024 [PDF]
« Our review shows that there is little support for any major influence of climate, neither by coarse-grained patterns nor by detailed spatiotemporal and mechanistic evidence. Conversely, there is increasing, strong support for human pressures as the key driver of these extinctions, with emerging evidence for an initial onset linked to pre-sapiens hominins prior to the Late Pleistocene. »
From & Meiri, Big, flightless, insular and dead: Characterising the extinct birds of the Quaternary, Journal of biogeography, 2021 [Abstract]
« Our findings suggest that extinct bird species differed from extant birds by being larger, mostly restricted to islands, and often flightless. These factors made them especially vulnerable to human prosecution and to other anthropogenically related declines. Our modern understanding of birds is skewed with respect to the nature of avian faunas that existed before the current wave of human-induced extinctions changed our world forever.«
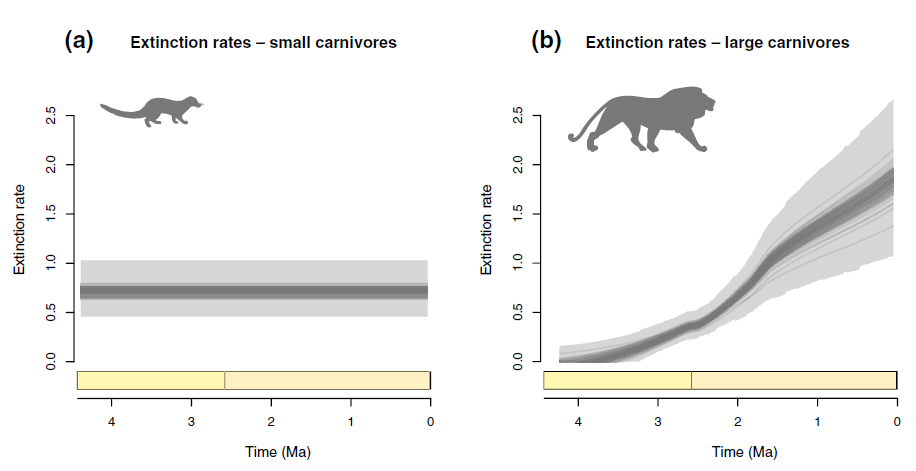
Faurby et al., Brain expansion in early hominins predicts carnivore extinctions in East Africa, Ecology letters, 2020 [Texte]
« While the anthropogenic impact on ecosystems today is evident, it remains unclear if the detrimental effect of hominins on co‐occurring biodiversity is a recent phenomenon or has also been the pattern for earlier hominin species. We test this using the East African carnivore fossil record. We analyse the diversity of carnivores over the last four million years and investigate whether any decline is related to an increase in hominin cognitive capacity, vegetation changes or climatic changes. We find that extinction rates in large carnivores correlate with increased hominin brain size and with vegetation changes, but not with precipitation or temperature changes. While temporal analyses cannot distinguish between the effects of vegetation changes and hominins, we show through spatial analyses of contemporary carnivores in Africa that only hominin causation is plausible. Our results suggest that substantial anthropogenic influence on biodiversity started millions of years earlier than currently assumed.«
Note : cette étude constate en Afrique une accélération du rythme des extinctions de grands prédateurs autour de -2 millions d’années, bien corrélée avec la présence d’hominines et l’accroissement de leur cerveau, ce qui laisse penser que la concurrence des hominines aurait pu être redoutable dès cette période, soit par kleptoparasitisme (voir mon article sur le charognage), soit par la chasse. Elle est contestée notamment par Faith et al, The uncertain case of human-driven extinctions prior to Homo sapiens, Quaternary Research, 2020.
Wißing et al., Stable isotopes reveal patterns of diet and mobility in the last Neandertals and first modern humans in Europe, Nature Scientific Reports, 2019 [Texte]
« Using the stable isotopic approach, we shed light on aspects of diet and mobility of the late Neandertals and UPMHs from the cave sites of the Troisième caverne of Goyet and Spy in Belgium. We demonstrate that their diet was essentially similar, relying on the same terrestrial herbivores, whereas mobility strategies indicate considerable differences between Neandertal groups, as well as in comparison to UPMHs. Our results indicate that UPMHs exploited their environment to a greater extent than Neandertals and support the hypothesis that UPMHs had a substantial impact not only on the population dynamics of large mammals but also on the whole structure of the ecosystem since their initial arrival in Europe.«
Smith et al., Body size downgrading of mammals over the late Quaternary, Science, 2018 [PDF]
« Here, we quantify mammalian extinction selectivity, continental body size distributions, and taxonomic diversity over five time periods spanning the past 125,000 years and stretching approximately 200 years into the future. We demonstrate that size-selective extinction was already under way in the oldest interval and occurred on all continents, within all trophic modes, and across all time intervals. Moreover, the degree of selectivity was unprecedented in 65 million years of mammalian evolution. The distinctive selectivity signature implicates hominin activity as a primary driver of taxonomic losses and ecosystem homogenization. Because megafauna have a disproportionate influence on ecosystem structure and function, past and present body size downgrading is reshaping Earth’s biosphere.«
Price et al., Big data little help in megafauna mysteries, Nature, 2018 [Texte]
« But in our view, the ‘big-data’ approach cannot, at this point, get us closer to an answer. There simply aren’t enough good-quality data. An understanding of what drove the extinctions requires detailed analysis on a species-by-species basis. This means investing effort into finding more fossil specimens and verifying the ages of those that have already been discovered using improved dating methods. It also means relating the timing of species’ existence and disappearance to detailed local environmental, climatic and archaeological records. »
L’incertitude des datations, les lacunes dans les données archéologiques rendent incertaines les méta-analyses. Notons que pour la précision des datations, des progrès majeurs ont été réalisés ces dernières années, mais les méta-analyses continuent d’utiliser aussi de vieilles données incertaines :
On trouvera une traduction en français dans cet article du blog Le Partage.
Carotenuto et al., The well-behaved killer: Late Pleistocene humans in Eurasia were significantly associated with living megafauna only. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018
« Although now-extinct megafauna was available to humans, it was rare where humans were most abundant. Medium sized, living megafauna shows the strongest association with humans. For Late Pleistocene megafauna in Eurasia, human hunting may have been just an additional, non-decisive extinction factor.«
Haynes, The Evidence for Human Agency in the Late Pleistocene Megafaunal Extinctions, Encyclopedia of the anthropocene, 2018 [Abstract]
« But the extinctions at the end of the Pleistocene are uniquely different because they unfolded almost instantly on an evolutionary timescale and had a disproportionate bias for megafauna, a term once applied to any animal larger than a rabbit and now meaning animals with average adult body mass44 or 45 kg (100 lbs). Alroy (2001, p. 1893) deemed the Late Pleistocene extinctions “unparalleled in the deeper fossil record” of North America, which they also seem to be on the other continents. This set of extinctions is not just another example of natural processes eliminating many animal genera; it is a first sign of irreversible human effects on the biosphere.«
Zhu et al., The large mean body size of mammalian herbivores explains the productivity paradox during the Last Glacial Maximum, Nature ecology and evolution, 2018 [Texte]
« The present-day results of potential grazer biomass, combined with an empirical land-use map, infer a reduction in wild grazer biomass by 79–93% owing to anthro-pogenic land replacement of natural grasslands. For the LGM, we find that the larger mean body size of mammalian herbivores than today is the crucial clue to explain the productivity paradox, due to a more efficient exploitation of grass production by grazers with a large body size. »
Lima-Ribeiro & Diniz-Filho, Climate change, human overkill, and the extinction of megafauna: a macroecological approach based on pattern-oriented modelling, Evolutionnary ecology research, 2017 [PDF]
« such overkill scenarios unrealistically predicted earlier extinction times than that observed for Megatherium. Moreover, the high human population growth required to simulate overkill scenarios was attained only recently after the industrial revolution, specifically in the mid-1900s, and it is therefore not applicable for Pleistocene hunter-gatherers. Conversely, although less frequent across simulations, models that correctly predicted the observed extinction time (16%) revealed plausible and empirically acceptable demographic parameters, encompassing low growth rates (rh<0.70%) and mortality for the human population(mo<0.60%), combined with moderate values of individual meat consumption (CI>70 g) andgeographical range collapse in Megatherium, which is produced by climate change. Conclusions: These findings, based on POM reasoning, highlight that unique mechanisms such as the overkill explanation for LQE, although feasible from model simulations, only occur with implausible parameter combinations and predict unrealistic extinction dynamics. »
Abramson et al., On the roles of hunting and habitat size on the extinction of megafauna, Quaternary international, 2017 [Texte]
« We show that the presence of hunters drives the superior herbivore to extinction even in habitats that would allow coexistence, and even when the pressure of hunting is lower than on the inferior one. The role of system size and fluctuating populations is addressed, showing an ecological meltdown in small systems in the presence of humans. The time to extinction as a function of the system size, as calculated with the model, shows a good agreement with paleontological data. Other findings show the intricate play of the anthropic and environmental factors thatmay have caused the extinction of megafauna.«
Michael Dennis Cherney, Records of Growth and Weaning in Fossil Proboscidean Tusks as Tests of Pleistocene Extinction Mechanisms, University of Michigan Library, 2016 [PDF]
« Tusk analyses for Ziegler Reservoir mastodons (Snowmass Village, CO) show no evidence that populations were struggling during the previous interglacial (Sangamonian) when climate was similar to current conditions. Poor nutrition is likely to result in later weaning age in mammals. However, in the interval of warming leading up to their extinction, Siberian woolly mammoths were apparently weaning earlier than they had been during the last glacial maximum. The shift to earlier weaning at the end of the Pleistocene refutes climate-related nutritional stress as a mechanism for their extinction. Population pressure from human hunting, which is expected to result in earlier weaning, is a more likely explanation for mammoth population declines. »
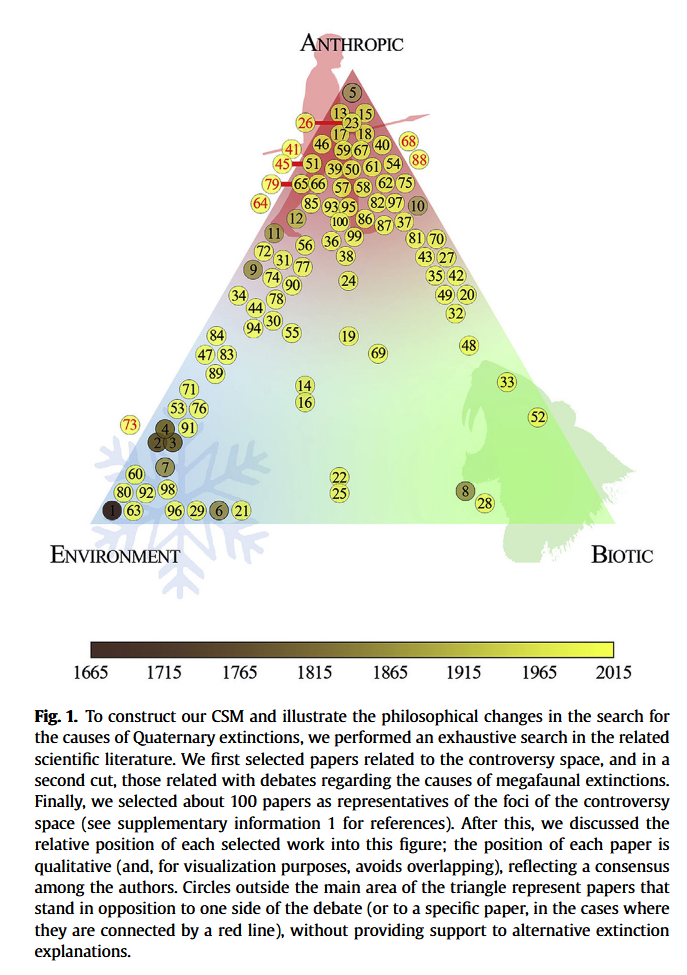
Monjeau et al., The controversy space on Quaternary megafaunal extinctions, Quaternary international, 2015 [PDF]
« We postulate that nowadays this controversyspace is suffering a period of conceptual blockage. This may be because authors are assembled around two major paradigms: environmental versus anthropic causes. Each of these two theoretical positionslooks at a portion of reality that may be partially true, but incomplete in terms of a global theory ofextinction. We propose that this conceptual blockage could be solved by developing a mathematicalmodel in which each hypothesis plays a role in a mechanistic way. The relative importance of eachhypothesis may vary depending on its respective context. It follows from this that it should not matterwhich cause is favored: the emphasis should be given to all causes acting together in a predictablemanner. »
Note : Revue systématique des publications sur la question des extinctions. La majorité des auteurs penche pour une cause plutôt anthropique, bien que peu de publications concluent à un rôle unique de l’action humaine.
Bartlett et al., Robustness despite uncertainty: regional climate data reveal the dominant role of humans in explaining global extinctions of Late Quaternary megafauna, Ecography, 2015 [PDF]
« Our results show that human colonisation was the dominant driver of megafaunal extinction across the world but that climatic factors were also important. We identify the geographic regions where future research is likely to have the most impact, with our models reliably predicting extinctions across most of the world, with the notable exception of mainland Asia where we fail to explain the apparently low rate of extinction found in in the fossil record. Our results are highly robust to uncertainties in the palaeological record, and our main conclusions are unlikely to change qualitatively following minor improvements or changes in the dates of extinctions and human colonisation.«
Pires et al., Pleistocene megafaunal interaction networks became more vulnerable after human arrival, Proceedings of the royal society, 2015 [PDF]
« We show Pleistocene and modern assemblages share similar network topology, but differences in richness and body size distributions made Pleistocene communities significantly more vulnerable to the effects of human arrival. The structural changes promoted by humans in Pleistocene networks woul dhave increased the likelihood of unstable dynamics, which may favour extinction cascades in communities facing extrinsic perturbations.«
Cooper et al., Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover, Science, 2015 [PDF]
« The causes of the Pleistocene extinctions of large numbers of megafaunal species in the Northern Hemisphere remain unclear. A range of evidence points to human hunting, climate change, or a combination of both. Using ancient DNA and detailed paleoclimate data, Cooper et al. report a close relationship between Pleistocene megafaunal extinction events and rapid warming events at the start of interstadial periods. Their analysis strengthens the case for climate change as the key driver of megafaunal extinctions, with human impacts playing a secondary role. »
Anthony John Stuart, Late Quaternary megafaunal extinctions on the continents: a short review, Geological journal, 2014 [PDF]
« This paper provides an overview of the contentious issue of global megafaunal extinctions in the Late Quaternary. The main proposed causes are ‘overkill’, environmental change or a combination of both. There are major objections to the other suggested causes. Extinctions were highly variable in their severity between different zoogeographical regions, with the greatest impact in North America, South America and Australia, but also substantial in northern Eurasia. Sub‐Saharan Africa and Southern Asia were much less affected. For northern Eurasia, detailed chronologies show a staggered extinction pattern, in which each megafaunal species exhibits unique and complex distributional shifts, culminating in extinction for some species and survival in others. Environmental drivers were clearly very important, although the possible role of humans is not yet clear. Alaska/Yukon also has a good radiocarbon record which also suggests a staggered extinction pattern. However, the available data for the rest of North America are largely unsatisfactory. South America also boasted spectacular extinct megafauna, but again the currently available dates are insufficient to reliably discern patterns or possible causes. Australia and New Guinea also suffered major losses, but extinctions probably occurred much earlier than elsewhere, so that establishing a chronology is especially difficult. Africa and Southern Asia have the least available data. In order to make meaningful progress, it is vital to establish a large database of reliable radiocarbon dates for each region made directly on securely identified megafaunal remains. The need is for much more high quality data, not more debate based on imperfect evidence. »
Sandom et al., Global late Quaternary megafauna extinctions linked to humans, not climate change, Proceedings of the royal society B, 2014 [Texte]
« We present, to our knowledge, the first global analysis of this extinction based on comprehensive country-level data on the geographical distribution of all large mammal species (more than or equal to 10 kg) that have gone globally or continentally extinct between the beginning of the Last Interglacial at 132 000 years BP and the late Holocene 1000 years BP, testing the relativeroles played by glacial–interglacial climate change and humans. We show that the severity of extinction is strongly tied to hominin palaeo biogeography, with at most a weak, Eurasia-specific link to climate change. This first species-level macroscale analysis at relatively high geographical resolution provides strong support for modern humans as the primary driver of the worldwide megafauna losses during the late Quaternary. »
Nogués-Bravo et al., Climate Change, Humans, and the Extinction of the Woolly Mammoth, PLOS Biology, 2008 [PDF]
« Results of the population models also show that the collapse of the climatic niche of the mammoth caused a significant drop in their population size, making woolly mammoths more vulnerable to the increasing hunting pressure from human populations. The coincidence of the disappearance of climatically suitable areas for woolly mammoths and the increase in anthropogenic impacts in the Holocene, the coup de grâce, likely set the place and time for the extinction of the woolly mammoth.«
Johnson, Determinants of loss of mammal species during the Late Quaternary ‘megafauna’ extinctions: life history and ecology, but not body size, Proceedings of the Royak Society B, 2002 [PDF]
« First, large size was not directly related to risk of extinction;rather, species with slow reproductive rates were at high risk regardless of their body size. This findingrejects the ‘blitzkrieg’ model of overkill, in which extinctions were completed during brief intervals ofselective hunting of large-bodied prey. Second, species that survived despite having low reproductive ratestypically occurred in closed habitats and many were arboreal or nocturnal. Such traits would have reducedtheir exposure to direct interaction with people. Therefore, although this analysis rejects blitzkrieg as ageneral scenario for the mammal megafauna extinctions, it is consistent with extinctions being due tointeraction with human populations.«
Norman Owen-Smith, Pleistocene extinctions: the pivotal role of megaherbivores, Cambridge university press, 1987 [Abstract]
The elimination of megaherbivores elsewhere in the world by human hunters at the end of the Pleistocene would have promoted reverse changes in vegetation. The conversion of the open parklike woodlands and mosaic grasslands typical of much of North America during the Pleistocene to the more uniform forests and prairie grasslands we find today could be a consequence. Such habitat changes would have been detrimental to the distribution and abundance of smaller herbivores dependent upon the nutrient-rich and spatially diverse vegetation created by megaherbivore impact. At the same time these species would have become more vulnerable to human predation.
Etudes locales
Adesanya Adeleye et al., On the timing of megafaunal extinction and associated floristic consequences in Australia through the lens of functional palaeoecology, Quaternary Science Reviews, 2023 [Texte]
« Dung fungi from marine core identify the timing of megafaunal extinction.
Megafaunal extinction occurred in southern Australia ∼43.5 ka. Human pressure primarily drove extinction. Plant trait and fossil pollen reveal vegetation changes associated with extinction. Declines in large-seeded and fruited plants accompanied and followed extinction.«
Dembitzer et al. (Ben-Dor, Barkai), Levantine overkill: 1.5 million years of hunting down the body size distribution, Quaternary science reviews, 2022 [Texte] [PDF]
« Mean hunted animal masses 10,500 years ago, were only 1.7% of those 1.5 million years ago. Neither body size at any period, nor size change from one layer to the next, were related to global temperature or to temperature changes. Throughout the Pleistocene, new human lineages hunted significantly smaller prey than the preceding ones. This suggests that humans extirpated megafauna throughout the Pleistocene, and when the largest species were depleted the next-largest were targeted. Technological advancements likely enabled subsequent human lineages to effectively hunt smaller prey replacing larger species that were hunted to extinction or until they became exceedingly rare. »
Murchie et al., Collapse of the mammoth-steppe in central Yukon as revealed by ancient environmental DNA, Nature communication, 2021 [Texte]
« We observe a substantial turnover in ecosystem composition between 13,500 and 10,000 calendar years ago with the rise of woody shrubs and the disappearance of the mammoth-steppe (steppe-tundra) ecosystem. We also identify a lingering signal of Equus sp. (North American horse) and Mammuthus primigenius (woolly mammoth) at multiple sites persisting thousands of years after their supposed extinction from the fossil record. »
Stewart et al., Climate change, not human population growth, correlates with Late Quaternary megafauna declines in North America, Nature communications, 2021 [Texte]
« Recently, a new Bayesian regression technique was developed that overcomes this problem—Radiocarbon-dated Event-Count (REC) Modelling. Here we employ REC models to test whether declines in North American megafauna species could be best explained by climate changes, increases in human population densities, or both, using the largest available database of megafauna and human radiocarbon dates. Our results suggest that there is currently no evidence for a persistent through-time relationship between human and megafauna population levels in North America. There is, however, evidence that decreases in global temperature correlated with megafauna population declines. »
Lord et al., Pre-extinction Demographic Stability and Genomic Signatures of Adaptation in the Woolly Rhinoceros, Current biology, 2020. [Texte]
« Demographic analysis suggests stable population size until close to extinction. No increased inbreeding or reduced genomic diversity coinciding with human arrival »
Seersholm et al., Rapid range shifts and megafaunal extinctions associated with late Pleistocene climate change, Nature communications, 2020 [Texte]
« Our findings suggest that climate change affected the local ecosystem in Texas over the Pleistocene-Holocene boundary, but climate change on its own may not explain the disappearance of the megafauna at the end of the Pleistocene. »
–
Pleistocene Mammal Communities and Their Extinction, in History of terrestrial mammals in South America (Livre)
Thomas Defler
Springer, 2018
https://link.springer.com/chapter/10.1007/978-3-319-98449-0_14
During the Pleistocene the diversity of mammals in South America became extremely elevated. It seems that hyperdiversity reached the highest known in the world. There has been nowhere else where so many (about 37) megamammals (weighing more than 1000 kg) were found, all of which became extinct, during the last 8000–9000 years ago. Another 44 species or so of large mammals weighing more than 45 kg and less than 1000 kg also became extinct. Of course ecological factors played a huge role in leveling this out-of-balance fauna, but the intriguing question has always been about the role that human beings had to cause these mammals to go extinct. Since we are unsure of the date of arrival of human beings (which might have begun as early as 40,000 years ago), it is difficult to put an exact date on the beginning of this new ecological pressure, but between 12,000 and 8000 years ago, the last extinctions occurred, when we know that humans were living widely throughout the continent.
–
Thriving or surviving? The isotopic record of the Wrangel Island woolly ammoth population
Arppe et al.
Quaternary Science Reviews, 2019
https://www.sciencedirect.com/science/article/pii/S0277379119301398?via%3Dihub#!
Scenarios related to water quality problems stemming from increased weathering, and a possibility of a catastrophic starvation event as a cause of, or contributing factor in their demise are discussed.
–
Disappearance of Icelandic walruses coincided with Norse settlement
Keighley et al.
Molecular biology and evolution, 2019
https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msz196/5564176
The high value of walrus products such as ivory on international markets likely led to intense hunting pressure, which – potentially exacerbated by a warming climate and volcanism – resulted in the extinction of walrus on Iceland. We show that commercial hunting, economic incentives and trade networks as early as the Viking Age were of sufficient scale and intensity to result in significant, irreversible ecological impacts on the marine environment. This is to one of the earliest examples of local extinction of a marine species following human arrival, during the very beginning of commercial marine exploitation.
–
Population reconstructions for humans and megafauna suggest mixed causes for North American Pleistocene extinctions
Broughton & Weitzel
Nature communications, 2018
https://www.nature.com/articles/s41467-018-07897-1
The results suggest that the causes for extinctions varied across taxa and by region. In three cases, extinctions appear linked to hunting, while in five others they are consistent with the ecological effects of climate change and in a final case, both hunting and climate change appear responsible.
–
Plio-Pleistocene decline of African megaherbivores: No evidence for ancient hominin impacts
Faith et al.
Science, 2018
https://science.sciencemag.org/content/362/6417/938.full
It has long been proposed that pre-modern hominin impacts drove extinctions and shaped the evolutionary history of Africa’s exceptionally diverse large mammal communities, but this hypothesis has yet to be rigorously tested. We analyzed eastern African herbivore communities spanning the past 7 million years—encompassing the entirety of hominin evolutionary history—to test the hypothesis that top-down impacts of tool-bearing, meat-eating hominins contributed to the demise of megaherbivores prior to the emergence of Homo sapiens. We document a steady, long-term decline of megaherbivores beginning ~4.6 million years ago, long before the appearance of hominin species capable of exerting top-down control of large mammal communities and predating evidence for hominin interactions with megaherbivore prey. Expansion of C4 grasslands can account for the loss of megaherbivore diversity.
–
The record of sloth coprolithes in north and south America : implications for terminal pleistocene extinctions.
Hunt & Lucas
New Mexico Museum of Natural History and Science Bulletin, 2018
https://www.researchgate.net/profile/Spencer_Lucas/publication/328676887_THE_RECORD_OF_SLOTH_COPROLITES_IN_NORTH_AND_SOUTH_AMERICA_IMPLICATIONS_FOR_TERMINAL_PLEISTOCENE_EXTINCTIONS/links/5bdb83004585150b2b98296f/THE-RECORD-OF-SLOTH-COPROLITES-IN-NORTH-AND-SOUTH-AMERICA-IMPLICATIONS-FOR-TERMINAL-PLEISTOCENE-EXTINCTIONS.pdf(lien non cliquable)
There is an acme of sloth coprolite preservation in the latest Pleistocene (also the only latrinites of Mammuthus and Bison) whichsuggests greater usage of caves by large herbivores at this time. The pattern of preservation of sloth coprolites suggests that climate rather than overkill was most significant in the extinction of ground sloths.
–
Chronological and Isotopic data support a revision for the timing of cave bear extinction in Mediterranean Europe
Terlato et al.
Historical biology, 2018
https://www.tandfonline.com/doi/abs/10.1080/08912963.2018.1448395?journalCode=ghbi20
The carbon (δ13C) and nitrogen (δ15N) isotopic values of bone collagen exhibit values similar to those of older cave bears from Swabian Jura and France, suggesting that the feedings preferences of cave bears remained unchanged until the disappearance of this species in Europe. Several bear remains preserved traces of human modification such as cut marks, which enables a reconstruction of the main steps of fur recovery and the butchering process.
–
Humans rather than climate the primary cause of Pleistocene megafaunal extinction in Australia
Van der Kaars et al.
Nature communications, 2017
https://www.nature.com/articles/ncomms14142
We record high levels of the dung fungus Sporormiella, a proxy for herbivore biomass, from 150,000 to 45,000 years ago, then a marked decline indicating megafaunal population collapse, from 45,000 to 43,100 years ago, placing the extinctions within 4,000 years of human dispersal across Australia. These findings rule out climate change, and implicate humans, as the primary extinction cause.
–
A simulation of anthropogenic Columbian mammoth (Mammuthus columbi) extinction
Matthew Clapman & Alex Capaldi
Historical Biology, 2017
https://www.researchgate.net/publication/320286050_A_simulation_of_anthropogenic_Columbian_mammoth_Mammuthus_columbi_extinction
Previous research has been conducted on the overkill hypothesis for the Columbian mammoth using a continuous differential equations model. We improved on this work by developing a computationally more efficient and more realistic discrete stochastic model. Most model parameters were obtained directly from the literature; migration parameters were informed by the literature and calibrated for the model. Our results provide evidence in support of the overkill hypothesis.
–
Au moins localement, les populations animales ont pu résister plus longtemps à la colonisation humaine.
At least 17,000 years of coexistence: Modern humans and megafauna at the Willandra Lakes, South-Eastern Australia
Westaway et al.
Quaternary Science Review, 2017
https://www.sciencedirect.com/science/article/abs/pii/S0277379116306011
We show that there is at least 17,000 years of overlap between people and megafauna at one of Australia’s richest archaeological landscape.
The research undermines the rapid kill hypothesis and suggests that megafauna extinction was likely the results of a combination of factors.
It is clear that much more research is required if we are to understand how the Australian megafauna became extinct.
–
Climate warming and humans played different roles in triggering Late Quaternary extinctions in east and west Eurasia
Xinru Wan & Zhibing Zhang
The royal society publishings, 2017
https://royalsocietypublishing.org/doi/10.1098/rspb.2016.2438
Here, our analyses showed that temperature change had significant effects on mammoth (genus Mammuthus), rhinoceros (Rhinocerotidae), horse (Equidae) and deer (Cervidae). Rapid global warming was the predominant factor driving the total extinction of mammoths and rhinos in frigid zones from the Late Pleistocene and Early Holocene. Humans showed significant, negative effects on extirpations of the four mammalian taxa, and were the predominant factor causing the extinction or major extirpations of rhinos and horses. Deer survived both rapid climate warming and extensive human impacts.
–
What caused extinction of the Pleistocene megafauna of Sahul?
Johnson et al.
Proceedings of the royal society B, 2016
https://royalsocietypublishing.org/doi/full/10.1098/rspb.2015.2399
During the Pleistocene, Australia and New Guinea supported a rich assemblage of large vertebrates. Why these animals disappeared has been debated for more than a century and remains controversial. Previous synthetic reviews of this problem have typically focused heavily on particular types of evidence, such as the dating of extinction and human arrival, and have frequently ignored uncertainties and biases that can lead to misinterpretation of this evidence. Here, we review diverse evidence bearing on this issue and conclude that, although many knowledge gaps remain, multiple independent lines of evidence point to direct human impact as the most likely cause of extinction.
–
Climate change not to blame for late Quaternary megafauna extinctions in Australia
Saltré et al.
Nature communications, 2016
https://www.nature.com/articles/ncomms10511/
When coupled with analysis of several high-resolution climate records, we show that megafaunal extinctions were broadly synchronous among genera and independent of climate aridity and variability in Australia over the last 120,000 years. Our results reject climate change as the primary driver of megafauna extinctions in the world’s most controversial context, and instead estimate that the megafauna disappeared Australia-wide ∼13,500 years after human arrival, with shorter periods of coexistence in some regions. This is the first comprehensive approach to incorporate uncertainty in fossil ages, extinction timing and climatology, to quantify mechanisms of prehistorical extinctions.
–
Synergistic roles of climate warming and human occupation in Patagonian megafaunal extinctions during the Last Deglaciation
Metcalf et al.
Science Advances, 2016
https://advances.sciencemag.org/content/2/6/e1501682
We identify a narrow megafaunal extinction phase 12,280 ± 110 years ago, some 1 to 3 thousand years after initial human presence in the area. Although humans arrived immediately prior to a cold phase, the Antarctic Cold Reversal stadial, megafaunal extinctions did not occur until the stadial finished and the subsequent warming phase commenced some 1 to 3 thousand years later. The increased resolution provided by the Patagonian material reveals that the sequence of climate and extinction events in North and South America were temporally inverted, but in both cases, megafaunal extinctions did not occur until human presence and climate warming coincided. Overall, metapopulation processes involving subpopulation connectivity on a continental scale appear to have been critical for megafaunal species survival of both climate change and human impacts.
–
Pleistocene Overkill and NorthAmerican Mammalian Extinctions
David J. Meltzer
Annual review of anthropology, 2015
https://pdfs.semanticscholar.org/a493/17a65367a412784e35b3e74f14ff11905f61.pdf
Overkill proponents argue that there is more archaeological evidence than we oughtto expect, that humans had the wherewithal to decimate what may have been millions of animals, and that the appearance of humans and the disappearance of the fauna is too striking to be a mere coincidence. Yet, there is less to these claims than meets the eye. Moreover, extinctions took place amid sweepingclimatic and environmental changes as the Pleistocene came to an end. It has long been difficult to link those changes to mammalian extinctions, but the advent of ancient DNA, coupled with high-resolution paleoecological, radiocarbon, and archeological records, should help disentangle the relative role of changing climates and people in mammalian extinctions.
–
Extinct New Zealand megafauna were not in decline before human colonization
Allentoft et al.
PNAS, 2014
https://www.pnas.org/content/pnas/111/13/4922.full.pdf
. Contradicting previous claims of a decline in moa before Polynesian settlement in New Zealand, our findings indicate that the populations were large and stable before suddenly disappearing. This interpretation is supported by approximate Bayesian computation analyses. Our analyses consolidate the disappearance of moa as the most rapid, human-facilitated megafauna extinction documented to date.
–
Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea)
Wroe et al.
PNAS, 2013
https://www.pnas.org/content/110/22/8777.short
Human involvement in the disappearance of some species remains possible but unproven. Mounting evidence points to the loss of most species before the peopling of Sahul (circa 50–45 ka) and a significant role for climate change in the disappearance of the continent’s megafauna.
–
Holocene survival of Late Pleistocene megafauna in China: a critical review of the evidence
Turvey et al.
Quaternary Science Reviews, 2013
https://www.sciencedirect.com/science/article/abs/pii/S0277379113002503?via%3Dihub
Evidence for Holocene survival of any Chinese Late Pleistocene megafaunal species is weak or untenable.
–
New perspectives on middle Pleistocene change in the large mammal faunas of East Africa: Damaliscus hypsodon sp. nov. (Mammalia, Artiodactyla) from Lainyamok, Kenya
Faith et al.
Paleogeography, Paleoclimatology, paleoecology, 2012
https://www.sciencedirect.com/science/article/pii/S0031018212004579?via%3Dihub
Although Lainyamok is no longer represented entirely by extant species, the absence of species common earlier in the middle Pleistocene of East Africa suggests substantial faunal turnover between 500 and 400 ka. Damaliscus hypsodon persisted in East Africa until the end of the Pleistocene and its extinction can be attributed to a loss of arid grassland environments at the onset of the Holocene. The fossil evidence from southern Kenya suggests that the development of the taxonomically modern large mammal community was a long-term process characterized by the extinction of grazing specialists, with marked turnover occurring between ~ 500 and 400 ka and near the end of the Pleistocene.
–
Pleistocene Megafaunal Collapse, Novel Plant Communities, and Enhanced Fire Regimes in North America
Gill et al.
Science, 2009
http://doc.rero.ch/record/210391/files/PAL_E4398.pdf
we established that the megafaunal decline closely precededenhanced fire regimes and the development of plant communities that have no modern analogs. The lossof keystone megaherbivores may thus have altered ecosystem structure and function by the release ofpalatable hardwoods from herbivory pressure and by fuel accumulation. Megafaunal populationscollapsed from 14,800 to 13,700 years ago, well before the final extinctions and during the Bølling-Allerød warm period. Human impacts remain plausible, but the decline predates Younger Dryas coolingand the extraterrestrial impact event proposed to have occurred 12,900 years ago
–
Characteristics of Pleistocene megafauna extinctions in Southeast Asia
Louys et al.
Palaeogeography, palaeoclimatology, palaeoecology, 2007
https://www.sciencedirect.com/science/article/abs/pii/S0031018206004032
The loss of these species cannot be assigned to a single cause. Rather their disappearance is likely tied to both climatic and human agents. Unlike other regions which experienced megafauna extinctions, eustatic changes in sea level in Southeast Asia seems to have been an important factor.
–
Rise and Fall of the Beringian Steppe Bison
Shapiro et al.
Science, 2004
https://science.sciencemag.org/content/306/5701/1561
The widespread extinctions of large mammals at the end of the Pleistocene epoch have often been attributed to the depredations of humans; here we present genetic evidence that questions this assumption. We used ancient DNA and Bayesian techniques to reconstruct a detailed genetic history of bison throughout the late Pleistocene and Holocene epochs. Our analyses depict a large diverse population living throughout Beringia until around 37,000 years
before the present, when the population’s genetic diversity began to decline dramatically. The timing of this decline correlates with environmental changes associated with the onset of the last glacial cycle, whereas archaeological evidence does not support the presence of large populations of humans in Eastern Beringia until more than 15,000 years later.
–
A Multispecies Overkill Simulation of the End-Pleistocene Megafaunal Mass Extinction
John Alroy
Science, 2001
http://pdfs.semanticscholar.org/169d/f12bcba2fe44d8e41c3fdc885944a7e24afa.pdf
A computer simulation of North American end-Pleistocene human and largeherbivore population dynamics correctly predicts the extinction or survival of32 out of 41 prey species. Slow human population growth rates, randomhunting, and low maximum hunting effort are assumed; additional parametersare based on published values. Predictions are close to observed values foroverall extinction rates, human population densities, game consumption rates,and the temporal overlap of humans and extinct species. Results are robust tovariation in unconstrained parameters. This fully mechanistic model accountsfor megafaunal extinction without invoking climate change and secondaryecological effects.
Extinction de Neandertal
Les données sur l’extinction de Neandertal devenant conséquentes, elles sont regroupées sur une page dédiée.
Controverses dans les médias et les blogs
Si l’on arrivait à la conclusion que les humains sont responsables d’extinctions depuis le paléolithique, quelles conclusions pourrait-on en tirer ? Existerait-il alors une nature humaine mauvaise, dont il faudrait conclure qu’elle serait incurable ?
Le mythe de l’homme tueur
Ana Minsky
Le Partage, 2019
https://www.partage-le.com/2018/11/le-mythe-de-lhomme-tueur-par-ana-minski/
D’après Martin, l’extermination de la mégafaune marque le début de la « Sixième extinction » qui se poursuit et s’accélère de nos jours. Sa théorie suggère que nous — Homo sapiens — sommes destructeurs et agressifs par nature, et certains n’hésitent pas à affirmer qu’il nous est impossible de lutter contre cette nature.
–
Non, l’humanité n’a pas toujours détruit l’environnement
Nicolas Casaux & Ana Minski
Reporterre, 2018
https://reporterre.net/Non-l-humanite-n-a-pas-toujours-detruit-l-environnement
De plus en plus d’individus hissés au rang de personnalités du mouvement écologiste par le système médiatique propagent une vision particulièrement réductrice et nuisible de l’espèce humaine. De l’astrophysicien Aurélien Barrau à Vincent Mignerot et son association Adrastia, l’idée selon laquelle l’être humain a toujours été un destructeur (« L’homme, ce tueur en série », ainsi que le titre Le Point) ne cesse de gagner en popularité. S’ils n’en tirent pas exactement les mêmes conclusions, il n’en reste pas moins que ce que cela suggère est problématique pour de multiples raisons.
–
Article du Monde se basant sur :
Body size downgrading of mammals over the late Quaternary
Smith et al.
Science, 2018
https://science.sciencemag.org/content/360/6386/310
L’homme, tueur en série des grands mammifères
Nathaniel Hertzberg
Le Monde, 2018
https://www.lemonde.fr/sciences/article/2018/04/23/l-homme-tueur-en-serie-des-grands-mammiferes_5289378_1650684.html
La cause ? Longtemps les scientifiques se sont opposés sur le sujet. Une météorite, comme lors de la disparition des dinosaures, il y a 66 millions d’années ? Un changement brutal du climat ? Ou encore l’homme, chasseur habile et sans scrupule ? Dans un article publié dans la revue Science, vendredi 20 avril, une équipe américaine vient appuyer cette dernière hypothèse. Elle n’accuse pas seulement les conquérants du Nouveau Monde, probablement arrivés par le détroit de Behring, d’avoir éradiqué quelques espèces imposantes. Elle nous juge responsables de « la baisse de la taille des mammifères à travers le quaternaire tardif », comme l’indique sobrement le titre de la publication. En d’autres termes, nous serions coupables de la disparition générale des plus grands mammifères sur les cinq continents, et cela depuis au moins 125 000 ans.
–
L’homme, ce tueur en série
Frédéric Lewino
Le Point, 2012
https://www.lepoint.fr/science/l-homme-ce-tueur-en-serie-19-07-2012-1696613_25.php
Que de cadavres dans le sillage d’Homo sapiens ! Sitôt qu’il met le pied sur un nouveau continent, le glas sonne pour de nombreuses espèces animales et humaines. Les mammouths en Asie et en Amérique, les ours des cavernes en Europe, le kangourou géant en Australie, mais aussi les hommes de Neandertal en Europe, de Denisova et de Florès en Asie. N’est-ce qu’une fâcheuse coïncidence ? La conséquence de changements climatiques simultanés ? Ou bien l’homme modernen’est-il qu’un effroyable tueur en série ? Les multiples massacres et génocides dont il s’est rendu coupable durant les temps historiques sont déjà un indice peu favorable sur son comportement durant les temps préhistoriques.
–
–
–
